 Evolution
Evolution
Seeing Ghosts in the Bushes — Or How to Keep the Theory of Evolution from Breaking Your Heart and Driving You Crazy
What would be evidence against evolution, and very strong evidence at that, would be the discovery of even a single fossil in the wrong geological stratum….But not a single solitary fossil has ever been found before it could have evolved.
Richard Dawkins (2009, pp. 146-7)
Professor Dawkins is right: you can’t be older than your own grandfather, all country-western songs notwithstanding.
But he is wrong about out-of-order fossils. As the recent excitement about the tetrapod trackways in Poland illustrates, the relationship between stratigraphic position — i.e., where in the geological column a fossil group occurs — and phylogenetic position — i.e., where cladistic analysis predicts the group should occur — is anything but straightforward. Indeed, the concept of a “ghost lineage” (see below) was introduced nearly 20 years ago by paleontologist Mark Norell, to accommodate the frequent lack of congruence between fossils and cladograms. Grandpa and grandma fossil are often too young, and grandchildren too old. (Actually these are mixed-up sister group, not ancestor-descendant, relationships, but I really wanted to use that Ray Stevens song link, and “I’m my own sister group” isn’t quite idiomatic.)
The puzzle is what to make of this lack of congruence, when it occurs. One might naively read Dawkins’s “evidence against evolution” statement as a genuine prediction, and then be mystified, or heartbroken, or driven crazy, when the theory of common descent motors right around anomalous data, such as the tetrapod trackways. Kinda does have the flavor of a country-western song: “You said you’d be testable and true / but you ran off when those fossils came through / oh evolution why’d I ever listen to you…”
Deeper insights await, however. Here is the moral, so you’ll know where the story is going:
Most evolutionary theorists — both those who study only living organisms, and those who study extinct (fossil) groups — understand that the theory of common descent must rule out certain states of affairs, to remain empirically testable. Hence, the hypothetical “Cambrian rabbit” that would explode common descent, if such a fossil were ever actually found in Cambrian strata.
Problem is, as systematist Gareth Nelson realized in 1978, “Cambrian rabbits” are found all the time (see Part 2 of this blog series). But these paleontological discoveries are not seen as challenging common descent, because that theory is tied up in a complex bundle with assumptions about — for instance — the completeness of the fossil record.
Thus, when fossils occur “out of evolutionary sequence,” what is ordinarily held as falsified, or tested, is not common descent at all, but something else entirely. Common descent doesn’t take the hit because the rest of the theoretical bundle absorbs the fossil anomalies.
If you think this means that common descent is not easily tested by paleontological data — well, you’d be right.
So, on to part 1 of the story. We should start with the concept of “ghost lineages.”
Why common descent needs ghosts in the bushes
Let’s begin with a basic logical point. Dawkins says that evolution — meaning the theory of common descent — would be tested, and falsified, if fossils were found out of evolutionary sequence, or “before they could have evolved,” as he puts it. This would be the case, however, only if we could predict evolutionary sequence independently of where fossils actually turn up in the geological record.
Enter cladistics. Using the anatomical characters of extant and fossil groups (and molecular data as well for extant groups), and assuming that the groups are related by common descent (remember that assumption, because we’ll come back to it later), we can generate a cladogram (a phylogenetic tree) specifying which groups should have branched first — i.e., as most primitive — and which later — i.e., as more, or most, derived. With that cladogram in hand, we can then turn to the fossil record to see if its sequence of first appearances fits with our cladistic prediction about the evolutionary relatedness of the groups in question.
Well, sometimes the fossil record cooperates: the branching order predicted by the cladogram matches the pattern of fossil first appearances (i.e., of representatives of the groups in question).
Often, however, the fossil record doesn’t fit the cladogram, a difficulty well known to paleontologists and systematists. In fact, the cladistic revolution in the 1970s and 80s was motivated in large measure by evolutionary theorists setting aside stratigraphy — the fossil record — as determining phylogeny: that is, in deciding who is related to whom, by what historical sequence. Writing in 1972, American Museum of Natural History paleontologists Bobb Schaeffer, Max Hecht, and Niles Eldredge bluntly expressed their skepticism about the relevance of the fossil record to phylogeny:
…the fossil record for most groups of organisms is too incomplete to allow the assumption that relative stratigraphic position is necessarily indicative of morphocline polarity. (1972, 37)
In simpler language: where a fossil occurs in the geological column may have nothing to do with its position in the evolutionary tree. Contra Dawkins, “derived” groups may be found too early, and “primitive” groups too late. The difficulty arises, as Schaeffer et al. continue (1972, 43), from
the frequent discovery that a morphologically “primitive” taxon may occur quite high in the stratigraphic range of a group for which it is supposed to be ancestral. This problem has vexed most paleontologists at one time or another, particularly in relation to groups with a poor fossil record.
To see the problem, consider a pair of diagrams from vertebrate paleontologist Mark Norell, taken from the 1992 paper where he formally introduced the concept of “ghost lineages.” Figure 1 shows a cladogram for groups ABCD, with their predicted branching order: first A, then B, then C and D last. This figure is also “calibrated,” by which Norell means that it predicts the stratigraphic (fossil) branching order as well: A and B should be present in the fossil record at time 9, whereas sister groups C and D should be jointly present at time 6.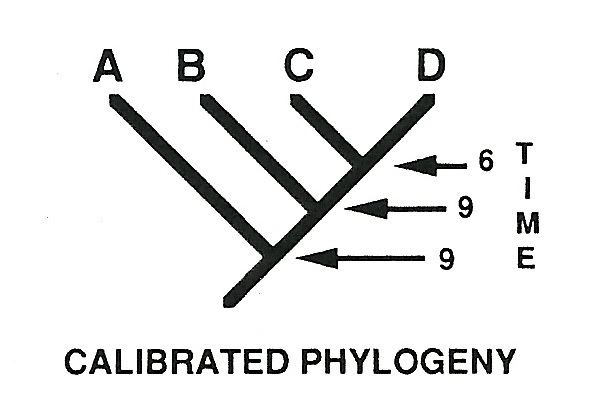
Figure 2, by contrast, shows the fossil (stratigraphic) distributions of groups ABCD. Note that A, the most primitive group, is found too late (“high in the range”), relative to its predicted branching order, whereas B is too early. Moreover, C and D fail to appear at the same time.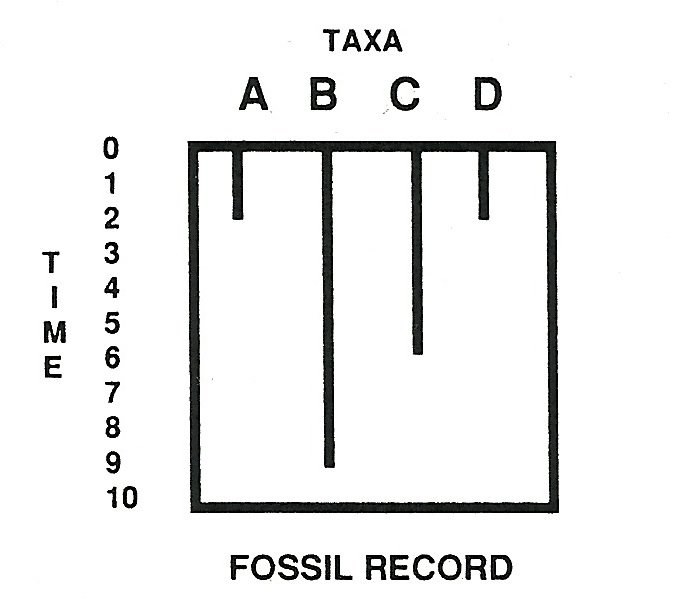
Now here we should pause, for a brief visit with logic. Read straightforwardly, the fossil record (in Norell’s toy example) has falsified the cladogram for groups ABCD. If the fossil record gives a reliable signal, groups ABCD simply could not be related in the order predicted by the cladogram: full stop. If you belong to group B, you can’t be older than group A, which supposedly branched earlier. Likewise, C and D could not be sister groups.
But hang on a minute. As noted above, the theory of common descent comes bundled with auxiliary theories and assumptions, among them assumptions about the completeness of the fossil record.
So — time to bring in the ghosts.
The role of ghost lineages in preserving hypotheses of common descent
Maybe group A just missed being preserved as fossils, but the group was actually present, albeit hidden, at time 9. Given the vagaries of fossilization, that’s a possibility. See Figure 3, then, where “ghost lineages” come to the rescue of the phylogeny ABCD.
The red circles are branching points, and the dotted red lines, ghost lineages.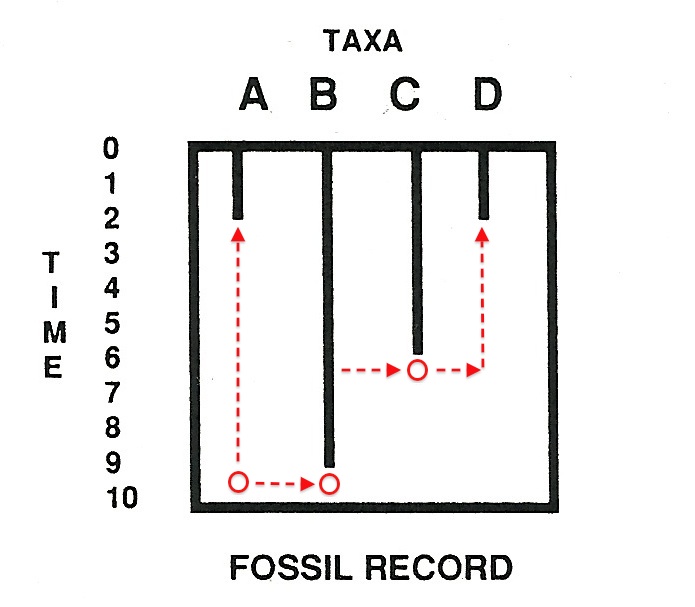
As Norell (1992, 105) writes,
These additional entities are taxa [groups] that are predicted to occur by the internal branching structure of phylogenetic trees….I refer to these as ghost lineages because they are invisible to the fossil record.
Ghost lineages — well, they just aren’t there, in any normal sense of “observable.” As Norell (1992, 105) notes, “ghost lineages are the unobservable predictions of the phylogenetic branching of the group ABCD” in the cladogram (Figure 1). It’s the cladogram, based on our assumption of common descent, which tells us the ghosts were there, and that groups A-D are related by common ancestry.
The fossils themselves don’t say that, however.
But, but — okay, slow down
At this point, haven’t we wandered a long way from Dawkins’s claim about the fossil record, cited at the top of this post? Isn’t common descent, rather than being tested by fossils, really providing instead the whole interpretative framework for the evidence — as an unquestioned first principle?
Yes — and that will be the theme of Part 2.
References
Dawkins, Richard. 2009. The Greatest Show on Earth: The Evidence for Evolution (New York: The Free Press).
Norell, Mark A. 1992. Taxic Origin and Temporal Diversity: The Effect of Phylogeny. In M.J. Novacek and Q.D. Wheeler, eds., Extinction and Phylogeny (New York: Columbia University Press), pp. 89-118.
Schaeffer, Bobb, Hecht, Max and Niles Eldredge. 1972. Phylogeny and Paleontology. Evolutionary Biology 6:31-46.