 Evolution
Evolution
 Life Sciences
Life Sciences
How to Play the Gene Evolution Game
| Links to our 8-Part Series, “The NCSE, Judge Jones, and Citation Bluffs About the Origin of New Functional Genetic Information”: • Part 1: Judge Jones’s Misguided NCSE-Scripted Kitzmiller Ruling and the Origin of New Functional Genetic Information Read the Full Article: “The NCSE, Judge Jones, and Citation Bluffs About the Origin of New Functional Genetic Information” |
The Gene Evolution Game is a very simple game to play. In three examples, we’ll develop three rules that can help you explain the origin of any new gene. That’s right–any gene! Let’s start with a simple example:
Rule 1: The Magic Wand of Gene Duplication
Where do new genes come from? Gene duplication is typically how we explain where a new gene comes from. Here’s how it works:
- (1) Take a gene you’ve observed in some organism. We’ll call it Gene B.
- (2) Find another gene similar to Gene B. Let’s call it Gene A.
- (3) Claim that at some time in the past, Gene A duplicated so then there were two copies of Gene A.
- (4) Then assert that one of Gene A’s duplicates evolved into Gene B.
Gene duplication is thus very a powerful explanation, and it looks like this:
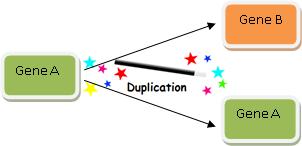
Wasn’t that easy? We’ve just explained how Gene B evolved! So when you find two genes with high sequence similarity, you can always explain how one evolved from the other via the magic wand of Gene Duplication.

The NCSE says “Gene duplication are [sic] common events, resulting from small errors in the process of cell replication. Once a gene is duplicated it is possible for one copy to mutate, adding information without risking the functioning of the pre-existing gene.” That’s all you need to know–when you invoke duplication, you needn’t worry about whether there is some functional evolutionary pathway for the duplicate gene to follow as it acquires some new function. In other words, you don’t need to worry about how new functional genetic information arises because “gene duplication” explains everything worth explaining! It’s easy to get extra genetic information in the Shannon sense, and that’s all that matters.
Rule 2: No Worries–Natural Selection Can Do It!
Now obviously the modern version of Gene B we find doesn’t perfectly resemble Gene A, or else it would be Gene A. So we have to account for how a copy of Gene A acquired its new function–Function B. One might think this would be the key part of explaining how new functional genetic information arises, but believe it or not, this is actually the easiest and quickest aspect of the game: we just call upon the power of “natural selection” and the problem is solved! This diagram shows exactly how we do it:
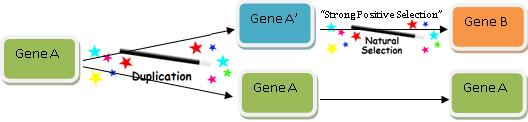
The great thing about the Gene Evolution Game is that natural selection can change (or not change, depending on what you wish) almost anything. And I mean anything.
Don’t worry about the details. If you want to account for differences between Gene B and Gene A, natural selection is always up to the challenge. Don’t worry about whether Gene A’ could evolve from Function A to Function B by small sequential adaptive steps. Don’t worry about the order in which amino acids changed, or whether many mutations were necessary to gain any functional advantage (that sort of thing is too unlikely to occur anyway, so just ignore it). Don’t worry about adaptive constraints, weak selection, or loss due to genetic drift. And most of all, definitely don’t do any calculations to determine the likelihood of whether all of the changes could have occurred in any reasonable amount of time.
We know the gene must have evolved, therefore it did evolve. Thus, you can think of natural selection as another magic wand. It may be invoked at any time to explain how a gene changed or evolved to acquire its new function.

This wand is a very powerful tool–it can explain both why things change, and why things stay the same.23 Wow!
Rule 3: The Magic Wand of “Rearrangement”
To play the Gene Evolution Game, there’s one last trick you need to know. Sometimes Gene B isn’t similar to just Gene A. Sometimes part of Gene B looks like Gene A, but another part looks like another gene. We’ll call the latter Gene Z. Don’t worry–this is all still easy to explain! We start by invoking duplication: Imagine that Gene A and Gene Z both duplicated, and then both duplicate copies were suddenly transported across the genome so that now they reside on a chromosome right next to one another. This is called “rearrangement.” If this sounds a little complicated, we’ll draw some diagrams to show how it works:
Step 1: Gene A and Gene Z are each in different locations, maybe even on different chromosomes:
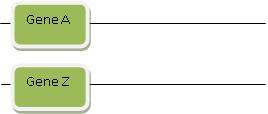
Then a special process called “rearrangement” suddenly rearranges and transports Gene A and Gene Z so they’re right next to each other in some other location in the genome. Rearrangement is a powerful magic wand you can invoke to explain how two stretches of DNA that initially are far apart suddenly end up near one another. They then can form a new functional gene. You’re probably getting a feel by now for how this works:

There are all kinds of rearrangements you can invoke–insertions, deletions, inversions, translocations–and you can invoke them in virtually any order and in any amount you please to explain how you get any two, or three, or even dozens of pieces of DNA to come together from throughout the genome to end up right next to one-another so that presto, you have your new functional gene. Just mix and match these types of rearrangements as needed to create virtually whatever DNA sequence you desire–rearrangement is always up to the task.

It’s all downhill from here. Natural selection can then perfect the rearranged gene to make it functional. Never mind detailed demonstrations that this actually works. Just sprinkle some natural selection and Gene A and Gene Z will magically combine functions and evolve into Gene B. Here we go, completing the explanation with everything we need to know:

Using the three magic wands of duplication, rearrangement, and natural selection you can provide a full and complete detailed explanation for the evolution of virtually any gene.
No Identifiable Ancestor? No Worries!
First, in some cases, your gene (i.e., Gene B) only has a homologue known from an entirely different species. So how did Gene B arrive in your organism? In these cases, just invoke lateral gene transfer (LGT) to whoosh the right gene into your organism. It doesn’t even matter whether lateral gene transfer is thought to occur between the organisms you’re working with–if the gene you need is found in some other species, then that by itself is evidence that lateral gene transfer occurs between the organisms you’re working with!24
Second, sometimes part of your gene doesn’t resemble part of any other known gene anywhere. Some people might wonder, “Where did this gene come from?” You still don’t worry about this. Remember what we said about natural selection? It can change anything. So if you can’t find any similar genes, just assume that your unique DNA sequence has evolved so much due to natural selection that it just doesn’t resemble its ancestral sequence any longer. But don’t worry, it’s not, and never is, the case that there wasn’t an ancestor. It’s just that the strong powers of natural selection changed the gene so much that we can’t identify any possible ancestral sequence.25
Some Final “Do’s” and “Don’ts” of the Gene Evolution Game
Right about now, you might be wondering about that last example we gave. So before you go any further, here’s a reminder of some questions you don’t need to ask:
- Given the known effects and rates of mutations, what were the odds of Gene A and Gene Z suddenly being rearranged next to one-another so that they could now function together as one single new gene product, Gene B?
- Did the rearranged gene product B start out functional? If not, how quickly could it gain function? How was it preserved from loss until it became functional?
- Are proteins really as malleable as this story would suppose or would the new combined gene encounter folding or other contextual problems?
- What mutational pathway was taken to evolve Gene A and Gene Z into a new gene with function B?
- What selective advantages were gained at each small step of this evolutionary pathway?
- Were any “large steps” (i.e., multiple specific mutations) ever required to gain a selective advantage along the evolutionary pathway? Would such “large steps” be likely to occur?
- Could all of this happen on a reasonable timescale?
You don’t need to worry about these questions. In fact, believe it or not, you don’t even need to know the function of your gene to claim it evolved from A and Z! All you need to know is that Genes A, Z, and B exist. This summary of these 3 simple rules of the Gene Evolution Game will help you explain anything:
- Gene Evolution Game Rule 1: Whenever you find sequence homology between two genes, just invoke a duplication event of some hypothetical, ancient ancestral gene, and you can explain how two different genes came to share their similarities.
- Gene Evolution Game Rule 2: When you need to explain how a gene acquired some new function, or evolved differences from another gene, just invoke the magic wand of natural selection. No need to demonstrate that there is any benefit to the new gene, or that a step-wise path to adaptation exists. Finally, natural selection is especially useful when part of your gene appears unique–since natural selection can change anything, just conclude that natural selection changed your gene so much that it no longer resembles its ancestor.
- Gene Evolution Game Rule 3: When a gene seems to be composed of the parts of several genes, just invoke duplications and rearrangements of all the DNA sequences you need, so you can get them all together in the right place. If you need to delete parts of a gene, or invert them, or transpose to a new location, just invoke different types of rearrangements as often and as liberally as you wish, and ba-da-bing, you’ve got your new gene!
And remember, don’t ask those other hard questions. Just use these three rules and you can explain virtually anything. No details required!
References Cited:
[23.] For example, when the ratio of nonsynonymous (i.e. amino acid changing) to synonymous (i.e. non-amino acid changing) differences between Gene B and Gene A is high, we have can say that it must be natural selection at work because only strong selection pressure would preserve so many changes that change amino acid sequence. Incredibly, we can also say that when the same ratio is low (i.e. there are FEW amino acid replacements in a gene), that too shows that natural selection was at work, in this case in the form of stabilizing selection to conserve gene sequence. This approach was taken in Harmit S. Malik and Steven Henikoff, “Adaptive Evolution of Cid, a Centromere-Specific Histone in Drosophila,” Genetics, Vol. 157:1293–1298 (March 2001) and its discussion of the Cid gene in a subsequent post.
[24.] See for example Ulfar Bergthorsson, Keith L. Adams, Brendan Thomason, and Jeffrey D. Palmer, “Widespread horizontal transfer of mitochondrial genes in flowering plants,” Nature, Vol. 424:197-201 (July 10, 2003). See also Mark A. Ragan and Robert G. Beiko, “Lateral genetic transfer: open issues,” Philosophical Transactions of the Royal Society B, Vol. 364:2241-2251 (2009) (“topological discordance between a gene tree and a trusted reference tree is taken as a prima facie instance of LGT”).
[25.] For example, this explanation was invoked in Matthew E. Johnson, Luigi Viggiano, Jeffrey A. Bailey, Munah Abdul-Rauf, Graham Goodwin, Mariano Rocchi & Evan E. Eichler, “Positive selection of a gene family during the emergence of humans and African apes,” Nature, Vol. 413:514-519 (October 4, 2001).
NOTE: This post is part 5 in a 8-part series responding to the NCSE, Judge Jones, Ken Miller, and the rest of the evolution lobby on the origin of new genetic information. This post, however, is written tongue-in-cheek and should be taken as a parody of the arguments of neo-Darwinists as they attempt to explain the origin of new genetic information. A number of examples showing neo-Darwinists specifically make the mistakes cited in this post will be shown in subsequent posts.