 Life Sciences
Life Sciences
Seeing Ghosts in the Bushes (Part 2): How Is Common Descent Tested?

If that dictum looks like a bumper sticker, I apologize — but it’s true all the same. Most of the philosophy of science can be captured by a handful of bumper stickers. Anyway, keep the dictum in mind. In this second installment of the “Seeing Ghosts in the Bushes” blog series — part 1 is here — we’ll ask how the theory of common descent could be tested by fossils. The principle of “what evidence cannot question, evidence cannot support” will be our main guide.
1. Hey, Common Descent Never Pays When She Runs a Stop Sign
Recall that Richard Dawkins argued common descent would be overturned if fossils occurred out of evolutionary sequence. “Evolution could so easily be disproved,” he writes (2009, p. 147), “if just a single fossil turned up in the wrong date order.” This prediction follows a long tradition in evolutionary reasoning, reaching back to the early 20th century, in which the possibility of “Precambrian rabbits” has been regarded as a crucial test of the theory of common descent.
But, as I’ll explain below, “Precambrian rabbits” are found all the time. These fossil discoveries aren’t seen as telling against common descent, however, because the “incompleteness of the fossil record” absorbs the blow. In part 1, for example, I showed how the concept of “ghost lineages” allows evolutionary theorists to accommodate anomalous stratigraphic (fossil) distributions. In short, when fossils occur out of order — as either too early or too late — ghost lineages mend the damage by invisibly extending the temporal ranges of groups, well beyond the actual fossil data, to achieve congruence with expectations. In part 2, I’ll show how common descent is further insulated from paleontological challenges by a protective buffer of auxiliary hypotheses.
Think about the problem this way. When a prediction of common descent fails, who pays for the failed prediction?
The cop writes a ticket for the broken prediction and hands it through the window. The vehicle belongs to common descent (CD), and indeed, she was driving when it was pulled over. Nevertheless, CD passes the ticket to one of her passengers.
“Take care of this for me, won’t you?” she says, as she sips her imported bottled water.
You get the picture. Common descent never pays for a ticket, because everyone else picks up the cost.
Let’s start with an influential paper by two leading paleontologists, from the early 1990s.
2. Norell and Novacek 1992: How Well Does the Fossil Record Match Cladistic Predictions?
In 1992, vertebrate paleontologist Mark Norell — who coined the term “ghost lineage” — and his American Museum of Natural History colleague Michael Novacek, asked an important question: How well does the fossil record fit with evolutionary expectations based on cladistic analysis? (A cladistic analysis of fossil and extant groups uses their characters to derive a cladogram, an evolutionary tree predicted independently of the groups’ stratigraphic [geological] position.) As Norell and Novacek explain,
Our purpose was to examine the reliability of the fossil record in recovering the sequence of fossil divergence events. The cladistic result [prediction] is not necessarily the closer match to the “true” phylogeny; it simply provides an independent means of assessing the timing of evolutionary events in the fossil record. (1992, p. 1691)
To assess the congruence of cladograms and fossils, Norell and Novacek assigned ranks to each. See Figure 1, from their paper: “Age rank” gives the location in geological time of the fossil distributions of groups ABCD, whereas “clade rank” gives the branching or phylogenetic order of the same groups, based in this case not on stratigraphy, but on the anatomical (or molecular) characters of ABCD.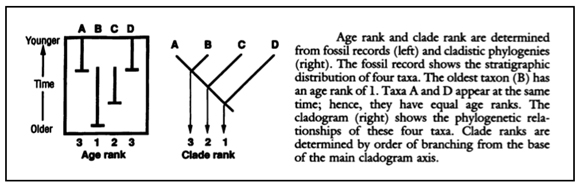
Fig. 1. Norell and Novacek (1992, p. 1692) assigned ranks to vertebrate groups, using both stratigraphic distribution and cladistic branching order. These diagrams show the ranks for hypothetical groups ABCD.
When plotted in this way, the two ranks — if congruent for any group — should produce points lying along a diagonal. “Complete congruence between the fossil record and the cladogram,” write Norell and Novacek, “in sequence of ranks and level of precision will produce a diagonal line of points intersecting the axes at their origin” (1992, p. 1691; emphasis added). See Figure 2, where the blue diagonal represents congruence. Points falling above that line mark groups arising too early in the fossil record, with respect to their clade rank, whereas points below the diagonal represent groups arising too late.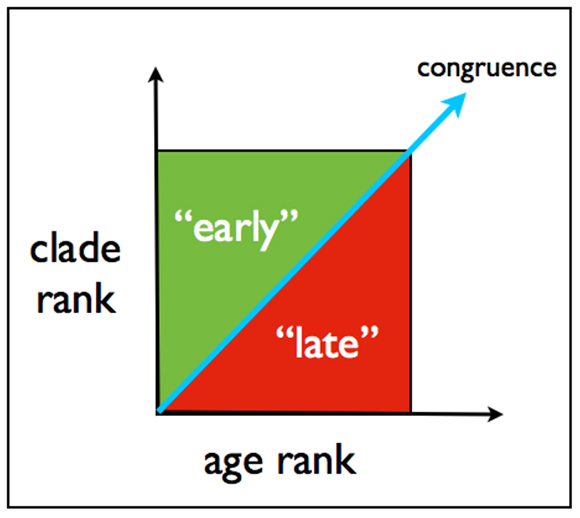
Fig. 2. For any taxonomic group, congruence between its age (fossil) rank and clade rank will produce a point lying on the blue diagonal. Points above that line represent groups that appear too early in the fossil record, with respect to their clade rank, whereas points below represent groups appearing too late in the fossil record with respect to clade rank.
Norell and Novacek then plotted age and clade ranks for several vertebrate groups. Figure 3 shows two such plots from their paper, for equids (i.e., horses and their relatives) and higher primates.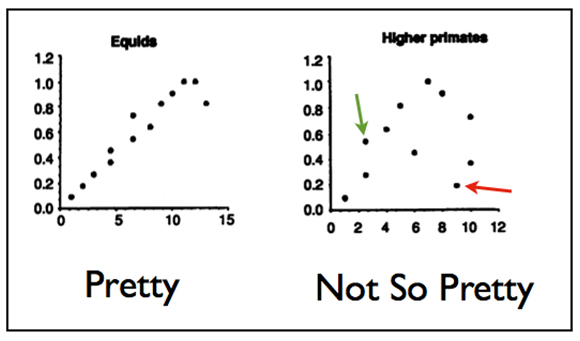
Fig. 3. Plots of age ranks as a function of clade ranks for two mammalian groups, equids and higher primates. The “pretty” plot for equids shows most of the points falling close to the diagonal, indicating (relatively) good congruence. The “not so pretty” plot for higher primates, by contrast, shows considerable scatter. For instance, the taxon marked by the green arrow appears too early in the fossil record, whereas the taxon marked by the red arrow appears too late.
Note that the equid plot corresponds well to the congruence diagonal, with only a couple of points straying. Norell and Novacek call this a “remarkable match of that [fossil] record with cladistic results” (1992, p. 1692). The higher primate plot, on the other hand, is frankly a mess. The taxon indicated by the green arrow, for instance, appears too early in the record, whereas the taxon marked by the red arrow appears very late (in both cases, with respect to cladistic predictions).
Now, before we look at how Norell and Novacek explained departures from theoretical expectation — including the not-so-pretty scatter in the higher primate plot — time for another visit with the logic of testing. We’ve got a handy bumper sticker for that.
3. When Data Don’t Cooperate
Testing a theory does not mean applauding whenever data and theory agree, while waving away or ignoring the instances where they don’t. That would be like assessing someone’s driving record by counting only the days when she didn’t speed or run through stop signs. Here’s another bumper sticker to make the point:
Remember the question motivating all this: How can common descent be challenged or questioned by paleontological evidence? Dawkins, following Julian Huxley, J.B.S. Haldane, and many other evolutionists, argues that common descent would be falsified if fossils were found out of evolutionary sequence.
This prediction may be represented schematically as follows (see Figure 4):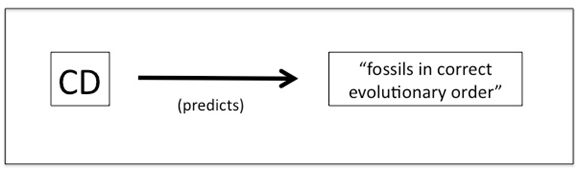
Fig. 4. Common Descent (CD) is widely said to predict that fossils should occur in the geological column in the correct evolutionary order.
If the prediction of “fossils in correct evolutionary order” follows from common descent, then when the prediction is overturned — as in the case of higher primate fossils (see the not-so-pretty plot, in Fig. 3, above) — common descent is falsified, at least for that group.
Right?
4. Don’t Be So Naïve, Paul
Of course not. Figure 5 shows what actually happens: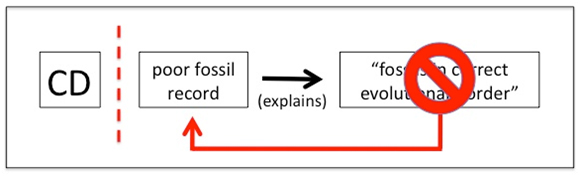
Fig. 5. When the prediction from CD of correct fossil order fails, the auxiliary hypothesis of “poor fossil record” absorbs the anomaly. CD is thereby protected from observational challenge.
Thus, when Norell and Novacek assessed the lack of congruence in the higher primate plot, guess who paid the ticket? Hint — not the driver:
The relationships examined here also reveal that the quality of the fossil record judged from other perspectives does not necessarily reveal its match with independently derived phylogenetic evidence….the primate fossil record poorly reflects higher level cladistic branching patterns. This is because some taxa (tarsiers and cheirogalines, for instance) thought to have branched off very early in primate history appear late in the record or have no fossil record. (1992, p. 1692; emphasis added)
Well, go figure. The higher primate fossil record is unreliable. Common descent is still true, of course. (In part 3 of this series, I’ll review a new theory for the common descent of the primates that sets their fossil record entirely to one side, as completely unreliable.)
In a deeply insightful paper published in 1978, which deserves to be better known, systematist Gareth Nelson grasped that what he calls “the paleontological argument” — namely, that fossils provide a reliable guide to evolutionary relationships — cannot be tested. Figure 5 shows why: the ad hoc hypothesis of the incompleteness of the fossil record absorbs any contrary evidence. As Nelson (1978, p. 331) writes,
Within the history of paleontology, I am aware of numerous apparent falsifiers of the paleontological argument; these have usually been rendered impotent as falsifiers by the ad hoc alternative that the fossil record was not as complete as previously believed….The paleontological argument seems fallacious because it is accepted in principle as non-falsifiable (it is always protected from falsification by an ad hoc alternative that is always, and obviously, true).
The “ad hoc alternative” is always true, because new fossils are always being found. As Nelson (1978, p. 331) notes — one can almost see him grinning — “The ad hoc alternative admittedly has appeal: how could its truth be doubted with the newly discovered fossils in hand?”
You can’t lose betting on the incompleteness of the fossil record. And, because you can’t lose, this ad hoc hypothesis works as a perfect sink to absorb any anomalous data the fossil record may reveal. Observe what happened recently with the discovery of the anomalously early Polish trackways. Did anyone question the common descent of the tetrapods?
Whaddya, crazy?
5. But No One Expects Common Descent to Pay for Her Tickets
This may seem a harsh judgment. It isn’t. Following the publication of Norell and Novacek 1992, a flurry of research began, led mainly by paleontologist Michael Benton and his co-workers, to examine the congruence of the fossil record and cladograms. I’ll discuss their research in greater depth in part 3. Here I want only to look at how Benton et al. handled the lack of congruence they found in one of their studies.
“What Causes Poor Matching of Age and Clade Data?” That’s the question Benton et al. asked in their 1999 paper on the problem (Benton, Hitchin and Wills 1999, p. 592). And here is their answer, which I have formatted as a series of bullet points for ease of reference:
Variations in congruence between cladograms and stratigraphic data result from several factors:
1. differences in the quality of cladograms;
2. differences in the quality of the fossil record;
3. stratigraphic problems;
4. categorical (taxonomic) focus; and
5. sampling density.
Only by looking at each case in some detail can we hope to determine the reasons for particularly good or particularly bad matching. (1999, p. 592)
Notice what theory is NOT in the list of possibilities to explain lack of congruence: the falsity of common descent itself. Figure 6 shows the logic. The investigator has (at least) five possible ad hoc or auxiliary hypotheses available to absorb any anomalous paleontological data.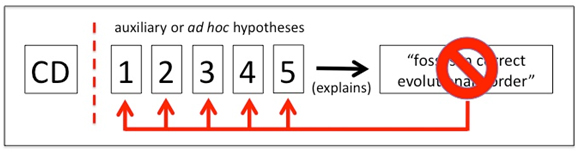
Fig. 6. Multiple possible ad hoc or auxiliary hypotheses are available to explain lack of congruence between the fossil record and cladistic predictions. These may be employed singly or in combination. Common descent (CD) is thus protected from observational challenge.
Common descent, although sitting in the driver’s seat, can pass any ticket she receives to one of her passengers. And she will.
Okay, I guess.
But what evidence cannot question, evidence cannot support.
In part 3, we’ll look at how all this plays out in current evolutionary research.
References
Benton, Michael, Rebecca Hitchin, and Matthew Wills. 1999. Assessing Congruence Between Cladistic and Stratigraphic Data. Systematic Biology 48:581-596.
Dawkins, Richard. 2009. The Greatest Show on Earth: The Evidence for Evolution. New York: The Free Press.
Nelson, Gareth. 1978. Ontogeny, Phylogeny, Paleontology, and the Biogenetic Law. Systematic Zoology 27:324-345.
Norell, Mark and Novacek, Michael. 1992. The Fossil Record and Evolution: Comparing Cladistic and Paleontologic Evidence for Vertebrate History. Science 255:1690-93.