 Evolution
Evolution
 Faith & Science
Faith & Science
 Life Sciences
Life Sciences
Discovering Signs in the Genome by Thinking Outside the BioLogos Box
Yesterday I promised that I would show you a mysterious genomic signal, and today I shall fulfill that promise. The previous blog was devoted to describing the linear distribution of LINEs and SINEs along mammalian chromosomal DNA. We saw that L1 retrotransposons tend to be densest in the regions where Alus and Alu-like elements are the least common and vice versa. I included the following figure from an article co-authored by Francis Collins1 that showed this compartmentalization of LINEs and SINEs along over a hundred million genetic letters of rat chromosome 10: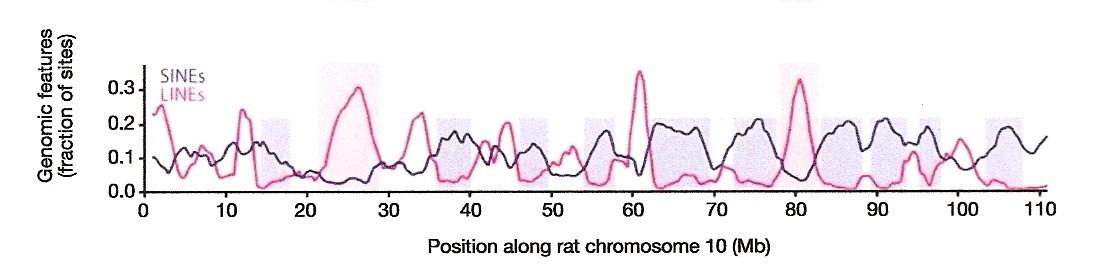
The blue line indicates the distribution of SINEs along a 110-million base pair interval of rat chromosome 10. (From Fig. 9d of Ref. 1.)
Taxon-Specific Elements: The SINEs Aren’t The Same
Intriguing as this non-random distribution of repetitive elements may be, it gets even more interesting when one realizes that SINEs are specific to taxonomic groups. Each primate genome has distinct subfamilies of the Alu sequence. The mouse genome, on the other hand, has no Alus but it does have three unique SINE families called B1, B2, and B4. While mouse B1 shares some sequence similarity with Alu, it has no relationship to the B2 or B4 elements; the latter two are also unrelated to each other. What then about the rat SINEs along chromosome 10, which were depicted as a blue line? Well, the genome of the rat has one main SINE family called ID, for the “Identifier” sequence. The ID elements have nothing in common at the DNA sequence level with the mouse B1s, B2s, or B4s, and they are wholly dissimilar to Alus.
What Would You Expect From Random Insertions Under A “Degenerative” Process?
So we have three different mammal genomes (primate, mouse, and rat) and three different sets of SINEs. But since I showed you rat chromosome 10 yesterday, let’s just focus on the two rodent genomes.
Now, the mouse and rat are estimated to have diverged twenty-two million years ago. During that interval, individual SINEs have been coming and going and going and coming, in and out of chromosomes. This ongoing insertion/deletion of these retrotransposons is precisely the “degenerative process” that Francisco Ayala referred to when mentioning Alus.
For the twenty-two million years that have occurred since the mouse and rat lineages went their separate ways, both genomes have been subjected to hundreds of thousands–if not millions–of separate SINE insertion events. Putting on our “junk DNA” thinking caps, let’s try to predict what the outcomes of such long-term mutational bombardments would be vis-à-vis the linear distributions of SINEs along a chromosome. To do this, let’s connect these two statements:
1) “…almost certainly much, if not most, of the DNA plays no role…”
2) “Perhaps one could attribute the obnoxious presence of the Alu sequences to degenerative biological processes…”
Or to restate, we have “much, if not most” rodent DNA that is not functional having being subjected to extensive degenerative events over the course of twenty-two million years. The only difference that we must keep in mind is that the “obnoxious” elements that were involved in this example of decay in the mouse genome are B1s, B2s, and B4s; whereas the destructive force in the rat genome in this case was primarily the ID elements.
Two Moons, Picking Up Impacts Independently — A Thought Experiment
Okay. What do we expect in general from degenerating processes that have no functional consequences? Let’s do a thought experiment. Consider the surfaces of two moons that were once part of the same planetary body 22 million years ago. Since their separation, both have been subjected to independent collisions with asteroids, meteorites, and other pieces of space debris. Question: Would you expect the scar patterns on both to be different or identical? (It may seem like a silly question, but bear with me.)
Replace now the word “moons” with the “mouse and rat genomes” and “asteroids and meteorites and other pieces of space debris” with SINEs, and you will see what I am asking. So I’ll rephrase my question. What should we expect regarding the linear distribution of independent SINE impacts along mouse and rat chromosomes?:
A. Completely independent patterns–like meteorite impact sites on moons;
B. A few overlapping patterns, due to chance; or
C. Nearly identical patterns.
And The Mystery Signal Is… 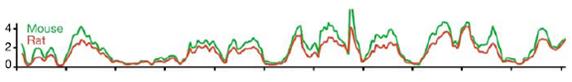
This is a second figure from the article co-authored by Francis Collins. (From Fig. 9c of Ref. 1.) The scale on the x-axis is the same as that of the previous graph–it is the same 110,000,000 genetic letters of rat chromosome 10. The scale on the y-axis is different, with the red line in this figure corresponding to the distribution of rat-specific SINEs in the rat genome (i.e., ID sequences). The green line in this figure, however, corresponds to the pattern of B1s, B2s, and B4s in the mouse genome.
Was it what you expected from a degenerative process? Why?
At this point the theistic evolutionist might say–Silly Rick: Common descent explains this pattern!
Wrong, wrong, wrong.
Let me repeat–each graph denotes only lineage-specific mutational insertions.
The mutational signal from mouse B1s, B2s, and B4s is equivalent to the mutational signal of rat IDs. It almost looks as if, say, the rat graph was copied, slightly redrawn, labeled “mouse,” and then pasted above the previous line. (Of course, it wasn’t.) How strange that two independently-acting degenerative processes–affecting mostly “junk DNA”– would lead to the same higher-order pattern.
It’s a bizarre pattern. And this correlation occurs throughout both genomes.
Not A Secret, Folks: Collins et al. Discussed This “Unusual” Finding
The Rat Genome Consortium–and thus Francis Collins–apparently thought it worthy to devote a whole section to the phenomenon. Titled Co-localization of SINEs in rat and mouse,1 the section states:
Despite the different fates of SINE families, the number of SINEs inserted after speciation in each lineage is remarkably similar: ~300,000 copies…Figure 9c displays the lineage-specific SINE densities on rat chromosome 10 and in the mouse orthologous blocks, showing a stronger correlation than any other feature. The cause of the unusual distribution patterns of SINEs, accumulating in gene-rich regions where other interspersed repeats are scarce, is apparently a conserved feature, independent of the primary sequence of the SINE and effective over regions smaller than isochores. (Italics mine.)
The potential signal in these two genomes, then, should be obvious. If not, I will belabor the point:
- The strongest correlation between mouse and rat genomes is SINE linear patterning.
- Though these SINE families have no sequence similarities, their placements are conserved.
- And they are concentrated in protein-coding genes.
Am I suggesting that extraterrestrials were fiddling with rodent DNA? No. Am I implying that we are seeing the “language of God” in rodent-script? I haven’t the foggiest notion. What I am saying is that we know a lot about the genome that is being glossed over in the popular works that the theistic evolutionists write. I am also saying that instead of finding nothing but disorder along our chromosomes, we are finding instead a high degree of order.
Is this an anomaly? No. As I’ll discuss later, we see a similar pattern when we compare the linear positioning of human Alus with mouse SINEs. Is there an explanation? Yes. But to discover it, you have to think outside the BioLogos box.
Reference
1. Rat Genome Sequencing Project Consortium. 2004. Genome sequence of the Brown Norway rat yields insights into mammalian evolution. Nature 428: 493-521.