 Evolution
Evolution
 Life Sciences
Life Sciences
PZ Myers, the Baldwin Effect, “Wolpert abuse,” and Quote-Mining
1. Call me a quote-miner, and I’ll see you outside — wait, you didn’t even read the article?
I’ll have a lot more to say about ontogenetic depth, in a “here’s where I think the idea needs to go” vein. This past weekend, however, PZ Myers responded to Parts I and II of the OD series, with what can only be described as a string of non-sequiturs.
That’s okay. Even non-sequiturs can provide useful teaching material. Myers’ response shows that natural selection does not solve the problem of building animals from single-celled or colonial ancestors, but I knew that already in 2004, when Myers and I talked at length at the Society for Developmental Biology meeting in Calgary. We can deal with the non-sequiturs later.
What is not okay, however, and requires an immediate reply, is Myers’ accusation that I quote-mined the evolutionary developmental biologist Lewis Wolpert. Quote-mining is the practice of citing scholarly sources out of context, for a polemical purpose. It’s a serious charge to make, and needs to be backed up with evidence.
Myers provides no such evidence, but that’s not the real problem. If you accuse someone of quote-mining, you actually have to read the source that supposedly was mined, to understand the context. If Myers had done so, he would have seen immediately that Wolpert makes exactly the same argument that I presented in the OD series: natural selection, at least as the process is conventionally modeled, cannot explain the origin of differentiation.
Myers has refused to say whether he read Wolpert 1994 before accusing me of quote mining the publication. As I’ll show below, the evidence indicates he did not read the article. Given that my argument and Wolpert’s are largely identical on the key point in question, Myers’ charge of quote-mining amounts to slander.
Let’s look at the evidence.
2. Why Invoke the Baldwin Effect? What IS the Baldwin Effect?
Richard Goldschmidt, one of the many who have attempted to envision a mechanism by which the complex epigenetic programs of metazoan development may have evolved, concludes:
If one tries to work out this idea in detail one soon comes to a point where it is evident that something besides the neo-Darwinian tenets is needed to explains such processes.
Given the neo-Darwinian tenet of the individual as the sole unit of selection, the dilemma is real and unresolvable.
— Leo Buss (1987, 117)
If one wants to simplify the problem of the origin of animals to bumper sticker length, one could put it this way, as a three-step puzzle:
1. Get cells to stick together.
2. Give them different jobs to do.
3. Repeat the whole cycle.
Because natural selection can only begin to operate after step 3, however (see part two of the OD series for a discussion of the necessary conditions of natural selection), preserving the variants that will cause steps 1-3 cannot be explained by selection. And indeed, to get his 1994 scenario going, Wolpert does not refer to natural selection at all:
Several changes are required in order for a single cell to give rise to a multicellular group by cell division…Firstly, the cell has to grow larger than its normal size and this requires a transient block to mitosis. Secondly, in the enlarged cell, the block over mitosis has to be released and the cell must then divide several times. And thirdly, the cells have to remain together….We can thus imagine a cell increasing its diameter 2.5-fold and then dividing four times to give 16 cells. Let us further assume that the third requirement is satisfied and the cells remain together, and, moreover, the cells form a hollow sphere. The latter may require oriented cell divisions or the maintenance of junctions between cells on the outer surface but that may not be too difficult. (1994, 80)
Figure 1, from Wolpert 1994, shows the sequence of events as Wolpert imagines it:
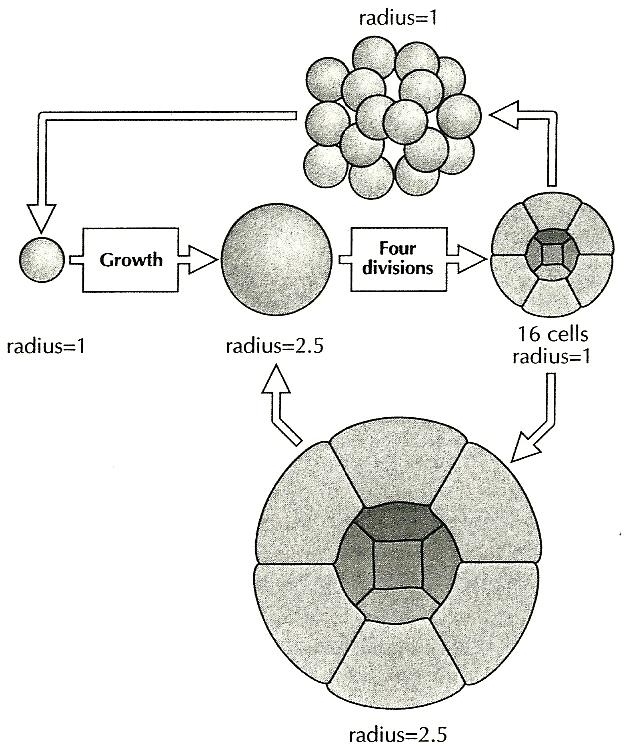
Figure 1. A single cell is assumed to grow without dividing. The cell then divides four times to form a hollow ball (Blastaea) of 16 cells. The process then repeats.
Okay — but what causal process does Wolpert see operating here, to bring about and preserve all of these changes?
The answer is the Baldwin effect: “an environmentally produced effect,” Wolpert argues, “becoming part of the developmental programme.”
If you’ve studied the history of evolutionary theory, chances are you’ve heard about the Baldwin effect. If you’ve studied pretty much the rest of modern biology — molecular, cell, developmental, systematic, etc. — chances are, you haven’t.
And that’s because the Baldwin effect is a hypothetical mechanism in search of supporting evidence. The central idea holds that phenotypic change causally precedes, or leads the way, to genotypic change; as West-Eberhard (2003) has put it, phenotypic accommodation leads to genetic accommodation. But does this happen in nature?
Massimo Pigliucci, next to West-Eberhard probably the leading theorist within evolutionary biology on the Baldwin effect and a related family of hypothetical processes, expresses caution (2010, 369-70):
The difficult task, of course, is to uncover convincing examples of transition from phenotypic to genetic accommodation and, moreover, to show that the phenomenon is sufficiently common to be evolutionarily relevant.
We’ll come back to that paucity of evidence. Let’s see first how Wolpert imagines the rest of his scenario to proceed.
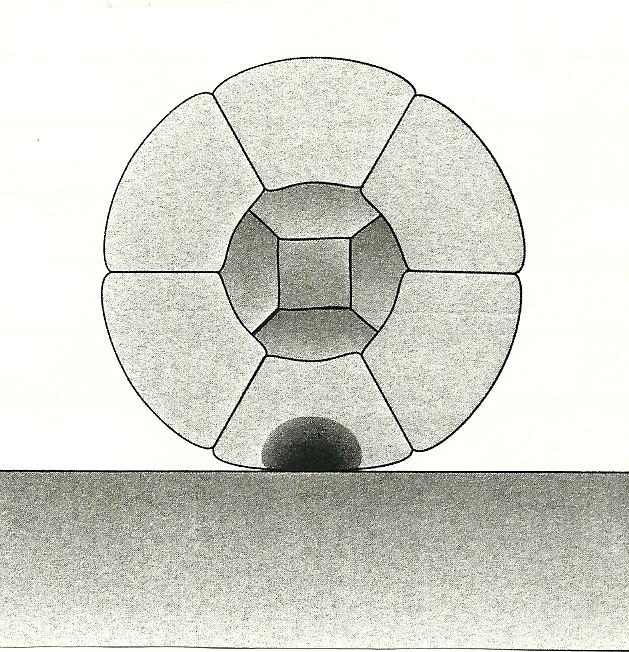
Figure 2 shows how Wolpert pictures the Baldwin effect as causing axes — polarized differences reflected in specialized cell populations — to evolve in an early embryo.
 ]
]
Figure 2. In Wolpert’s scenario, the Blastaea rests on a surface, where mechanical contact with that surface induces a permanent change in the lower-most cells, thus specifying an axis for the organism as a whole.
And Figure 3 shows how Wolpert envisions the Baldwin effect causing the origin of a germ line:
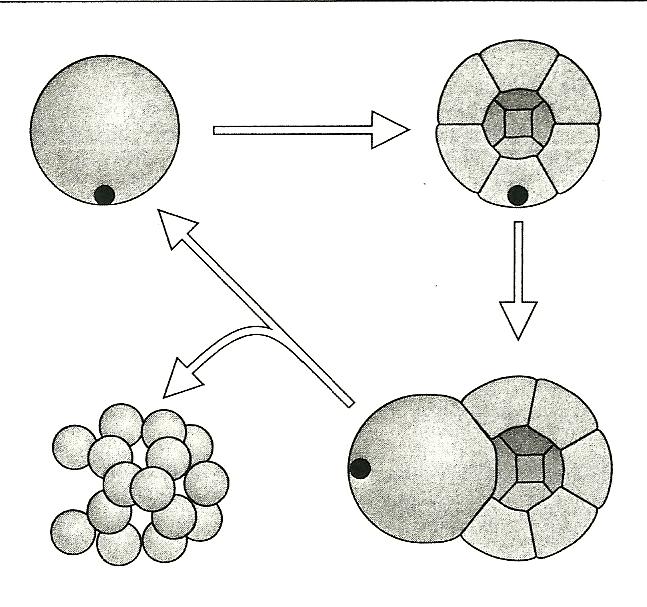
Figure 3. An unknown cytoplasmic factor is isolated by asymmetric cell division, causing only one cell to grow without dividing. This cell then becomes the germ line.
But why invoke the Baldwin effect at all? Why not model this series of events by starting with genetic mutations, which are then selected for their phenotypic effects? That is, why not describe the scenario as neo-Darwinian theory requires?
3. Because Natural Selection Wouldn’t Work
Wolpert explains that the Baldwin effect — the hypothetical direct action of external forces inducing permanent (stably heritable) cellular change — is needed to get around a causal problem with evolving differentiation de novo via natural selection:
In its absence [i.e., without the Baldwin effect] it would be necessary to first genetically specify one group of cells as being different but without there being any selective advantage. Only then could there be changes in these cells similar to those described above. This sequence of events is unlikely in the extreme. The reason for invoking the Baldwin effect is that an environmental signal provides an initial basis for a developmental alteration, which could have a selective advantage. (1994, 81; emphasis added)
Hey, sounds familiar, except for that Baldwin effect part — that is, except for the hypothetical evolutionary mechanism that is doing all the explanatory work in the story.
Let’s review Wolpert’s reasoning:
— In conventional neo-Darwinian theory, variations occur randomly and are selected for their effects. [This is standard theory.]
— However, with the origin of cellular differentiation — “one group of cells as being different,” in his words — the selective effect is downstream of reproduction, one of the necessary conditions of natural selection. [My point, in part II of the OD series, and the main point at issue.]
— Therefore, some process must cause permanently heritable variation prior to selection.
Enter the Baldwin Effect. Be sure to click on the link.
In the next entry, I look forward to analyzing PZ Myers’ cogent explanation for the origin of differentiation, which has elegant bumper sticker brevity:
{kind=link}
Step one is simply cell adhesion. Step two is gene regulation. Step three is epigenetics. That’s it.
Gosh, evolutionary theory is not as hard as I thought.
References
Buss, Leo. 1987. The Evolution of Individuality. Princeton University Press.
Pigliucci, M. 2010. Phenotypic Plasticity. In M. Pigliucci and G. M�ller, eds., Evolution: The Extended Synthesis. MIT Press.
West-Eberhard, M.J. 2003. Developmental Plasticity and Evolution. Oxford University Press.
Wolpert, L. 1994. The evolutionary origin of development: cycles, patterning, privilege and continuity. Development 1994 Supplement. 79-84.