 Evolution
Evolution
 Intelligent Design
Intelligent Design
The White Space in Evolutionary Thinking

When certain biologists discuss the early stages of life there is a tendency to think too vaguely. They see a biological wonder before them and they tell a story about how it might have come to be. They may even draw a picture to explain what they mean. Indeed, the story seems plausible enough, until you zoom in to look at the details. I don’t mean to demean the intelligence of these biologists. It’s just that it appears they haven’t considered things as completely as they should. Like a cartoon drawing, the basic idea is portrayed, but there is nothing but blank space where the profound detail of biological processes should be.
Let me give an example. This week Discovery Institute released a pair of videos (“How to Build a Worm” and “Switched on Worms“) featuring Fellow and philosopher of biology Paul Nelson and a lowly nematode called C. elegans. Its development is precise and intricately patterned, like a Bach fugue that splits and weaves many voices into one. The final cadence is the newly hatched larva. Not coincidentally, the videos use music by Bach throughout.
C. elegans‘s development is a wonder to behold, well worth sharing with the world. But there is more: the development of these lowly worms exposes a major problem with the Darwinian account of evolution.
The development of C. elegans is an end-directed process. As the videos reveal, each embryo follows a precisely choreographed developmental road map in order to get to the final goal — the reproductive adult. None of the intermediate steps on the way to the adult will do. Each step is necessary but not sufficient by itself. Turn aside from this developmental pathway and the result is likely to be a damaged worm or a dead one. Skip some steps and the same is true. How did this process come about? We would say this goal-directedness is evidence for a designer who had the final end in mind, and arranged the proper developmental steps appropriately.
Evolutionary biologists disagree. They say this exquisitely refined developmental pathway evolved gradually, a little at a time. First there was a cell, then a eukaryotic cell, one with a nucleus, organelles, and a cytoskeleton. Then along came multicellularity — cells living together to make an organism, with some cells set aside to make the next generation, and others free to specialize. As time went on, new digestive, muscle, nerve, and sensory cells evolved and were successfully coordinated into functioning whole organisms.
These organisms then adopted increasingly complex shapes and lifestyles (including the lifestyle of the nematode worm), and developed spicules, chitin, fur or feathers, the ability to creep or to fly, to see and to sing. All these things happened because of random genetic change and natural selection. No road map, no design was needed. How do we know this happened? Because we can find a series of animals that resemble each step in the story — Volvox, the sea urchin embryo, the planula of jellyfish, and flatworms like planaria. We find fossils that tell the story as more hard-bodied animals came along. The genes of these animals also speak of a common history (though the relationships are disputed for the deep histories). Finally, the series progresses logically, and purely by naturalistic means.
I should add that no one thinks Volvox, sea urchins, or flatworms (etc.) actually are ancestral to C. elegans or anything else. Rather these forms represent hypothetical “intermediates” that are discussed in a general way in textbooks (this may have happened, could have been, probably was…). Actual intermediates from unicellular eukaryotes to C. elegans are nowhere to be found, though. They are long gone, if they ever existed, so no one knows what they would have looked like. Despite this, names derived from these hypotheticals are assigned to clades, all based on genetic comparisons between extant species.
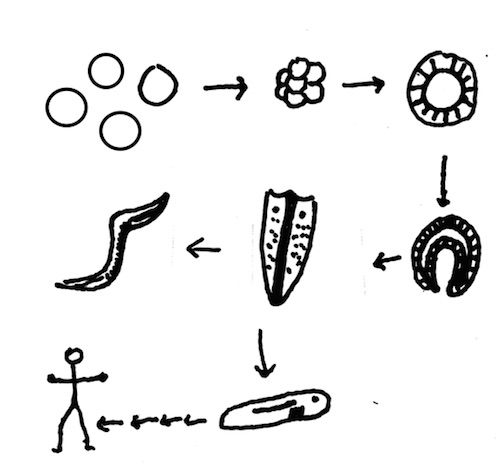
There’s a problem, though. The story leaves out too much. To make this clear I have drawn a cartoon version (there are other versions, of course, because nobody really knows how it happened).
-
A cell. That step’s a doozy (top left).
-
Cells stick together (like sponges?).
-
Cells form a hollow sphere with some set aside as reproductive cells, others for swimming and taking in food (like Volvox?).
-
Gastrulation. Cells move inward to form distinct layers and begin to differentiate one from another (like the sea urchin embryo?).
-
Another big step, a planula larva with bilateral symmetry, epithelium (a sheet of cells that encloses the body), a mouth, and perhaps a gut (like the planula of jellyfish? Not shown).
Over evolutionary time some planulae develop into something with a gut, epithelial, neural, muscular, and sensory cells (shown here as something like a planarium — a small flatworm with eyespots, nerves, epithelia, muscle, and a one-way gut). Other planulae become the animals we call jellyfish or comb jellies. Then comes the evolution of all other animal forms as documented by the fossil record. Two products are shown: C. elegans and, by way of an early chordate with a notochord and gill slits, us.
This cartoon of the evolutionary story is stripped down to its bare essentials. Does it seem oversimplified? It is. Besides the fact that no one knows what happened, in biology nothing is so simple. Imagining a series of animals in such a story does not take into account or explain what is necessary for transitions within the series to take place. Similarity, or a progression in forms, does not mean there is an undirected pathway between those things. The details of how the transitions took place must be supplied, and their reasonableness assessed.
New genes and proteins must be invented. The cytoskeleton, Hox genes, desmosomes, cell adhesion molecules, growth factors, microtubules, microfilaments, neurotransmitters, whatever it takes to get cells to stick together, form different shapes, specialize, and communicate must all come from somewhere. Regulatory proteins and RNAs must be made to control the expression in time and space of these new proteins so that they all work together with existing pathways.
In fact, in order for development to proceed in any organism, a whole cascade of coordinated genetic and biochemical events is necessary so that cells divide, change shape, migrate, and finally differentiate into many cell types, all in the right sequence at the right time and place. These cascades and the resulting cell divisions, shape changes, etc., are mutually interdependent. Interrupting one disrupts the others. In C. elegans changing one cell division can have significant effects on the fates of downstream cells, and the earlier the change, the more downstream effects there will be. Coordinated, finely tuned cascades of events that lead to functional organisms cannot be produced by chance and natural selection alone.
If all these processes happen in the right way — the road map is followed — the cells of C. elegans successfully divide to make a worm that swims, eats, and reproduces, helping to turn compost into soil. If other creatures follow their own roadmaps, chicks hatch and caterpillars turn into butterflies, kittens are born, and babies sleep in their mother’s arms.
According to evolutionary theory there is no roadmap, no design for how we got here. But after taking a closer look and filling in the details, it becomes apparent that there is a deep, coordinated, planned order in the world, as evidenced by C. elegans’s development or the development of any creature — an order with beauty and purpose, like a Bach fugue.