 Evolution
Evolution
 Intelligent Design
Intelligent Design
#5 of Our Top Stories of 2015: C. elegans and the Problem of Evolving Animal Body Plans
Editor’s note: Welcome to the traditional recounting of our Top 10 evolution-related stories of the past year, as compiled in a rigorous, peer-reviewed, strictly scientific manner by Evolution News staff. Sit back and enjoy the most amusing, the most enlightening, and the most important news to come our way in 2015. The countdown will culminate on New Year’s Day.
Happy New Year from your friends in the intelligent design community! If you haven’t finalized your year-end contribution to support the work of the Center for Science & Culture, including Evolution News, please do so now. Any amount helps. We need you and greatly appreciate your generosity!
![]()
Published originally on May 15, 2015.
Nearly all the animals we know have bilateral symmetry at some stage of their lives, meaning they have right and left halves that are mirror images of each other. These animals also have a head and a tail, a top and a bottom to them. The technical terms are anterior/posterior (A/P) and dorsal/ventral (D/V) axes (plural for axis, not the wood-chopping instrument). The exceptions are things like sponges, jellyfish, sea anemones, and small creatures like the Volvox I wrote about last week.
Even simple little worms like C. elegans have A/P and D/V axes, but how those axes get specified during the worm’s development is far from simple. It involves something like this pathway:
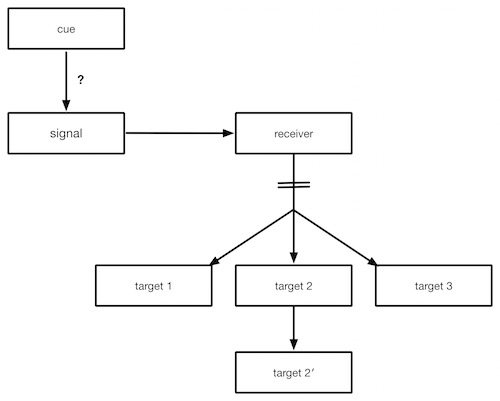
I have generalized the diagram to describe all signaling pathways. The specific details vary from organism to organism, from situation to situation. In C. elegans, subject of the new Discovery Institute video “How to Build a Worm,” the cue is the entry of the sperm into the egg at one end. This sets off the signal — in C. elegans‘s case it is the formation of an asymmetric arrangement of microtubules — structural elements involved in cell division. The asymmetric arrangement of microtubules causes a wave of cortical contractions that redistributes a protein in the cytoplasm to the cortex. (The cortex is the area of cytoplasm just under the cell membrane, and it is rich in structural proteins that cause movement and in developmental receptor molecules.) That protein then causes changes to other proteins embedded in the cortex. The asymmetrical redistribution of these proteins is represented as the receiver in the diagram. Then an asymmetrical cell division caused by the asymmetric microtubules partitions the proteins into different cells. This division is indicated by the slash marks in the diagram.
The worm has now established its A/P axis. The partitioned proteins go on to affect downstream targets in different ways. How the target molecules are affected influences the fates of all the future cells resulting from those first two cells, including which will become dorsal, and which ventral cells. If you want more detail, you can read about it in Scott Gilbert’s textbook, Developmental Biology.
The development of all bilaterally symmetric animals involves signaling networks. A signal is sent based on some cue, it is received, and some change is effected, over and over again. We see the developmental cascade that results in “How to Build a Worm.”
As shown there, the cascade of signaling leads to the production of multiple cell types arranged in a particular pattern that is C. elegans. The details differ from organism to organism — when the cells divide, when the signals are sent — but all bilaterally symmetric animals use something like the same process to set up their basic body plans and what cell types are made.
Even when the signaling pathway is stripped to its simplest, it still involves a cue to cause a signal to be sent and received. The cue is of no use unless it triggers a signal that can be received and cause downstream effects. That is what is necessary to begin to establish a difference in cell fates, and that requires a minimum of three molecules all to be present in order to work. It would take a very long time to assemble such a pathway, but would the result be of any selective advantage? Not unless there were more pathways involved that caused movement in the direction of some resource, or assigned a special function to particular cells that benefitted the whole.
These signaling pathways pre-date the animals that use them now. Based on genomic analyses, these signaling molecules have been around well before the first bilaterian animals ever existed. They are expressed in organisms that lack these body axes completely. Even more surprising, many of the molecules used to make complex structures such as muscles, eyes, and brains also predate their use for those purposes.
What were these signaling molecules used for before there was a left and right, a top and bottom, a head and tail? How did they come to be at all, and why did they persist until they could be co-opted for the establishment of body axes? It has been suggested that they were used to establish body sections in the earliest multicellular animals, but that only pushes the question back a step. Where did that use come from? How did the signaling pathways start?
With these questions, I conclude the series I began to address the white space in evolutionary thinking — how to account for the evolution of C. elegans. First there is the problem of getting a cell, then of getting a eukaryotic cell, then of getting a multicellular animal, and now of getting one with a head and a tail and multiple cell types. Saying C. elegans didn’t have to solve the problem all at once is merely to suggest that the problems are easier if taken one step at a time. They are not.