 Intelligent Design
Intelligent Design
From Genome to Body Plan: A Mystery
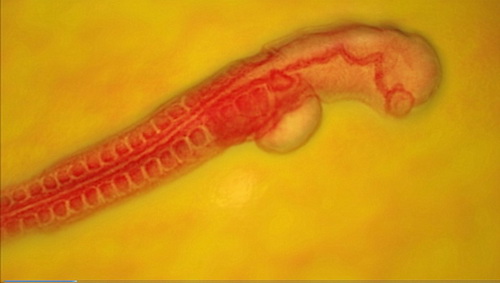
Decoding genomes has been one of the most important advances of the last sixty years, but it’s really just a start of a far larger mystery: the mystery of development. You can appreciate the magnitude of the problem in Illustra’s animation of a chick embryo in “Embryonic Development” from Flight: The Genius of Birds. An even more majestic depiction closer to home takes you from the moment of conception to the birth of a baby in this animation by RenderingCG. How does a linear genome produce such an astounding product? Then, how does the moving, living being reduce its information back down to a genome in a single cell?
Three German scientists discuss the mystery in a paper in Nature, “From morphogen to morphogenesis and back,” which can be loosely translated, “From genome to body plan and back.”
A long-term aim of the life sciences is to understand how organismal shape is encoded by the genome. An important challenge is to identify mechanistic links between the genes that control cell-fate decisions and the cellular machines that generate shape, therefore closing the gap between genotype and phenotype. The logic and mechanisms that integrate these different levels of shape control are beginning to be described, and recently discovered mechanisms of cross-talk and feedback are beginning to explain the remarkable robustness of organ assembly. The ‘full-circle’ understanding of morphogenesis that is emerging, besides solving a key puzzle in biology, provides a mechanistic framework for future approaches to tissue engineering.
Stop right there. Why must the framework be mechanistic? Didn’t they just speak of “the logic and mechanisms that integrate” at different levels? Logic is not mechanistic; it is conceptual. Logic can be instantiated in circuits, on paper, and in human language. Mechanism may be the primary aspect of morphogenesis that natural science can investigate, but restricting one’s investigation to a “mechanistic framework” is sure to miss the message in a book by considering only the paper and the ink.
After a brief history of morphogenesis theory from Aristotle to the era of molecular genetics, the authors claim that problems in the “mechanics centered approach” were finally solved in the 1970s. Here, they confuse the football players for the strategy of the play (so to speak). They describe the actions of the players, as if they operate mechanically, while hiding the quarterback’s game plan behind passive-voice verbs (“is controlled” — by whom?).
The initial landmark publication from this herculean project revealed that the first step in morphogenesis is the subdivision of the embryo into discrete regions by a cascade of ‘patterning’ genes4. Only then is each domain converted to the corresponding region of the body through a bespoke morphogenetic program, therefore establishing that the timing, positioning and inheritance of tissue-shaping events is controlled genetically. Subsequent molecular characterization in Drosophila and other systems revealed that patterning genes mainly encode signalling pathways that mediate long-range tissue patterning and gene-regulatory networks that control fate decisions; however, such genes do not control cell and tissue shape directly. Rather, the task of physically shaping cells and tissues is performed using a toolbox of essential cellular machines discovered by cell biologists, which are present in all cells in the embryo.
We appreciate the mention of a program, a toolbox, and machines, but who wrote the program? Who designed the tools and machines? It’s as if the authors are watching tools moving and operating without any hands:
Collectively, these studies reveal a picture in which the shape of tissues is determined by the combined actions of genetic, cellular and mechanical inputs (Box 1). Although a number of the main players are now known, and their functions understood, we still know surprisingly little about how the various levels of shape control are integrated during morphogenesis.
“Are integrated” — by whom? Passive voice verbs screen these authors from identifying plausible causes. And so by restricting their attention to how pieces of matter “are integrated,” they witness rabbits coming out of hats without a magician:
The focus of this Review is the logic and mechanisms that connect gene regulation, cellular effectors and tissue-scale mechanics — the troika of tissue shaping. We describe how shape, at the local level, emerges from the interaction of tissue-specific genetic inputs and the self-organizing behaviour of core intracellular machines. We then discuss how this mechanistic logic is used in several modified forms to produce a variety of shaping modes. It is becoming clear that the chain of command from gene to shape is not unidirectional, owing to the discovery of mechanisms that enable changes in tissue architecture and mechanics to feed back to ‘upstream’ patterning networks. The emerging integrated view of tissue shaping therefore goes full circle, from morphogen to morphogenesis and back.
Mechanistic philosophy gets hopelessly muddled here. To see why, convert the passive voice to active voice. “Mechanistic logic is used” should mean, “Somebody or something uses logic to operate a machine.” A baby’s shape doesn’t just “emerge” by “self-organizing behavior” except in the imagination of a philosophical materialist.
From there, the authors get into the weeds, discussing blastocysts, fruit flies, “evolutionarily conserved mechanosensitive pathways” and other matters. It should be obvious, though, that if you start on the wrong track you are not going to get where you want to go (i.e., understanding morphogenesis). In this dreamland, rabbits will pop out of hats by emergence. Babies will self-organize. Programs will work without a programmer.
The authors marvel at how “organoids” emerge from induced pluripotent stem cells. Is this an example of self-organization? After thinking about it, they admit that more must be going on.
A stunning demonstration of the full-circle nature of morphogenesis, in which genes regulate tissue shaping and vice versa, comes from the study of organoids. Here, cultured pluripotent cells self-assemble into organ-like structures that are remarkably similar to those formed in the embryo. Organoids can even be generated from patient-derived induced pluripotent stem cells, which means that this technology has the potential to herald a new era in tissue engineering for the modelling of disease and the development of therapies that is based on the principles of developmental biology…. Organoid formation itself demonstrates that cells can become organized in the absence of predetermined long-range external patterning influences such as morphogen gradients or mechanical forces, which are a cornerstone of classic developmental biology. This unexpected lack of requirement for long-range pre-patterning has led to organoid formation being described as an example of ‘self-organization’, which is defined classically as the spontaneous emergence of order through the interaction of initially homogeneous components. Although some aspects of organoid formation may show self-organizing properties, it is already clear that cell heterogeneity and patterned gene expression play a crucial part throughout.
The organoids will never form by self-organization, therefore, unless the coded instructions in each cell direct them according to “patterned gene expression” — that is what is crucial. They have a game plan, like band players in a “scatter” formation on the field self-organizing into a formation. Each player knows where to go.
The same issue of Nature takes a mechanistic look at the related issue of hierarchical organization. How does that “emerge”? In their article “Scaling single-cell genomics from phenomenology to mechanism,” Tanay and Regev begin:
Three of the most fundamental questions in biology are how individual cells differentiate to form tissues, how tissues function in a coordinated and flexible fashion and which gene regulatory mechanisms support these processes. Single-cell genomics is opening up new ways to tackle these questions by combining the comprehensive nature of genomics with the microscopic resolution that is required to describe complex multicellular systems. Initial single-cell genomic studies provided a remarkably rich phenomenology of heterogeneous cellular states, but transforming observational studies into models of dynamics and causal mechanisms in tissues poses fresh challenges and requires stronger integration of theoretical, computational and experimental frameworks.
Even though they seek a mechanistic framework again, they are employing intelligent design to get there: tackling questions, combining concepts, seeking causes. Will a “stronger integration of theoretical, computational and experimental frameworks” emerge by unguided material processes? Well, they seem to think cells did some remarkable things that way:
Multicellular organisms have evolved sophisticated strategies for cooperation between cells, such that a single genome encodes numerous specialized and complementary functional programs that maximize fitness when they work together. Compartmentalization at several levels — cells, tissues and organs — leads to functional diversification of cells and systems with the same underlying genome. Physical copies of the genome are embedded in cells to enable them to maintain a semi-autonomous decision-making process through the selective management of small-molecule, RNA and protein concentrations in cytoplasmic and nuclear compartments. Theoretically, this permits genomes to break the inherent symmetry that is imposed by the precise duplication of DNA in multicellular species. In particular, it facilitates cellular differentiation through the progressive acquisition of specific intracellular molecular compositions, enabling epigenetic mechanisms to emerge and implement cellular memory. At a higher level of organization, intercellular signalling, extracellular structures and environmental cues are used to form complex spatial structures in which cells (and their genomes) are physically embedded. This creates further levels of compartmentalization that encode complex and structured tissues.
More muddle. On the one hand, strategies, codes, programs, decision-making, cues, and signaling — implying rationality. On the other hand, evolution, emergence, and physical stuff — implying materialism. The authors mix oil and water, thinking the oil evolved out of the water and both cooked themselves into a soufflé.
After some diversion into issues like whether or not cell types can be classified in some Linnaean system, they take pride that science is beginning to move from descriptive accounts to predictive understanding:
Efforts towards the mapping and classification of cellular programs in humans and model organisms are becoming increasingly ambitious, aiming to provide a comprehensive atlas of the cell types and subtypes in organs and whole organisms. This opens up remarkable opportunities to move beyond descriptive studies of cell type and state and to develop mechanistic-predictive models of regulatory programs.
There’s no question that mechanisms are involved in development. But to mix in another metaphor, they’re focused on how billiard balls move and interact on the pool table but ignoring the expertise of the players. Even if the players are robots, and the shots are predictable and repeatable, you’ll miss the talent of the game without considering the intelligent design that directs each ball into its own pocket in the correct sequence. The design employs the laws of nature, but does not emerge from them.
Photo: Chick embryo, courtesy of Illustra Media.