 Evolution
Evolution
This Didn’t Evolve a Few Mutations at a Time
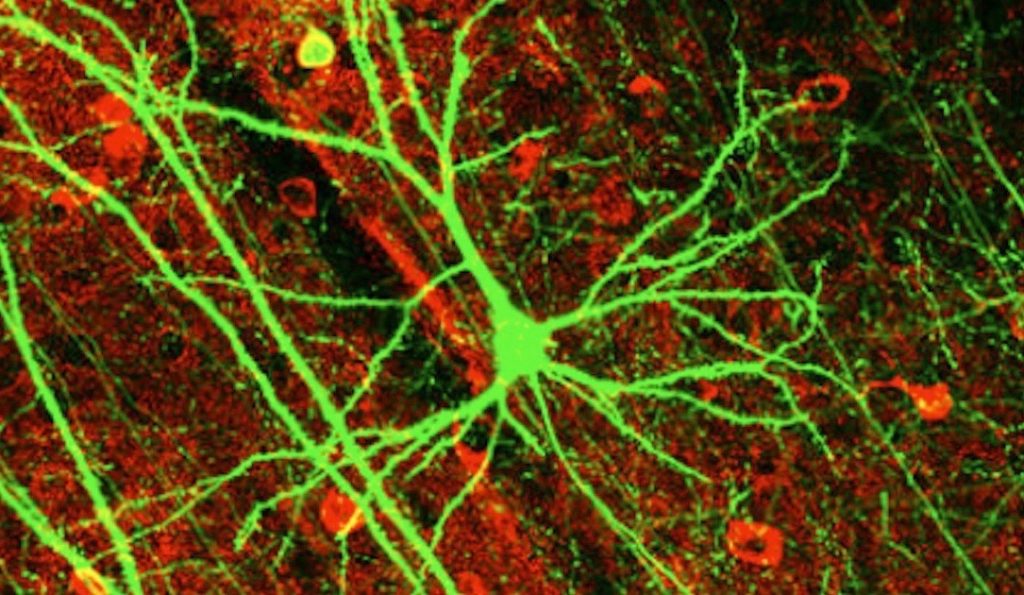
Are there long, gradual, pathways of functional intermediate structures, separated by only one or perhaps a few mutations, leading to every single species, and every single design and structure in all of biology? As we saw here yesterday, this has been a fundamental claim and expectation of evolutionary theory which is at odds with the science. Some evolutionists have attempted to get around the problem with the neutral theory, but that just makes matters worse.
If one mutation is rare, a lot of mutations are astronomically rare. For instance, if a particular mutation has a one-in-a-hundred million (one in 10^8) chance of occurring in a new individual, then a hundred such particular mutations have a one in 10^800 chance of occurring. It’s not going to happen. Let’s have a look at an example: nerve cells and their action potential signals.
Nerve cells have a long tail which carries an electronic impulse. The tail can be several feet long and its signal might stimulate a muscle to action, control a gland, or report a sensation to the brain.
Like a cable containing thousands of different telephone wires, nerve cells are often bundled together to form a nerve. Early researchers considered that perhaps the electronic impulse traveled along the nerve cell tail like electricity in a wire. But they soon realized that the signal in nerve cells is too weak to travel very far. The nerve cell would need to boost the signal along the way for it to travel along the tail.
After years of research it was discovered that the signal is boosted by membrane proteins. First, there is a membrane protein that simultaneously pumps two potassium ions into the cell and three sodium ions out of the cell. This sets up a chemical gradient across the membrane. There is more potassium inside the cell than outside, and there is more sodium outside than inside. Also, there are more negatively charged ions inside the cell so there is a voltage drop (50-100 millivolt) across the membrane.
In addition to the sodium-potassium pump, there are also sodium channels and potassium channels. These membrane proteins allow sodium and potassium, respectively, to pass through the membrane. They are normally closed, but when the decaying electronic impulse travels along the nerve cell tail, it causes the sodium channels to quickly open. Sodium ions outside the cell then come streaming into the cell down the electro-chemical gradient. As a result, the voltage drop is reversed and the decaying electronic impulse, which caused the sodium channels to open, is boosted as it continues on its way along the nerve cell tail.
When the voltage goes from negative to positive inside the cell, the sodium channels slowly close and the potassium channels open. Hence the sodium channels are open only momentarily, and now with the potassium channels open, the potassium ions concentrated inside the cell come streaming out down their electro-chemical gradient. As a result the original voltage drop is reestablished.
This process repeats itself as the electronic impulse travels along the tail of the nerve cell, until the impulse finally reaches the end of the nerve cell. Although we’ve left out many details, it should be obvious that the process depends on the intricate workings of the three membrane proteins. The sodium-potassium pump helps set up the electro-chemical gradient, the electronic impulse is strong enough to activate the sodium channel, and then the sodium and potassium channels open and close with precise timing.
How, for example, are the channels designed to be ion-selective? Sodium is about 40 percent smaller than potassium so the sodium channel can exclude potassium if it is just big enough for sodium. Random mutations must have struck on an amino acid sequence that would fold up just right to provide the right channel size.
The potassium channel, on the other hand is large enough for both potassium, and sodium, yet it is highly efficient. It somehow excludes sodium almost perfectly (the potassium to sodium ratio is about 10000), yet allows potassium to pass through almost as if there were nothing in the way.
Nerve cells are constantly firing off in your body. They control your eyes as you read these words, and they send back the images you see on this page to your brain. They, along with chemical signals, control a multitude of processes in our bodies, and there is no scientific reason to think they gradually evolved, one mutation at time.
Indeed, that idea contradicts everything we know from the science. And yet this is what evolutionists believe. Let me repeat that: evolutionists believe nerve cells and their action potential designs evolved one mutation at time. Indeed, evolutionists believe this is a proven fact, beyond all reasonable doubt.
It would be difficult to imagine a more absurd claim. So let’s have a look at the details of this line of thinking. Here is a recent paper from the Royal Society, representing the state of the art in evolutionary thinking on this topic. The paper claims to provide a detailed explanation of how early evolution produced action potential technology.
Sounds promising, but when evolutionists speak of “details,” they have something slightly different in mind. Here are several passages from the paper which reveal that not only is there a lack of details, but that the study is thoroughly unscientific.
We propose that the next step in the evolution of eukaryote DCS [membrane depolarization (through uncontrolled calcium influx), contraction and secretion] coupling has been the recruitment of stretch-sensitive calcium channels, which allow controlled influx of calcium upon mechanical stress before the actual damage occurs, and thus anticipate the effects of membrane rupture. [Emphasis added.]
The recruitment of calcium channels? And exactly who did the recruiting? Here the authors rely on vague terminology to paper over a host of problematic details of just how random mutations somehow performed this recruiting.
To prevent the actual rupture, the first role of mechanosensory Ca++ channels might have been to pre-activate components of the repair pathway in stretched membranes.
“To prevent”? Let’s spell out the logic a little more clearly. The authors are hypothesizing that these calcium channels evolved the ability to pre-activate the repair pathway “to prevent” actual rupture. By spelling out the logic a bit more clearly, we can see more easily the usual teleology at work. The evolution literature is full of teleology, and for good reason. Evolutionists are unable to formulate and express their ideas without it. The ever-present infinitive form is the tell-tale sign. Aristotelianism is dead — long live Aristotelianism.
As another anticipatory step, actomyosin might have been pre-positioned under the plasma membrane (hence the cortical actomyosin network detected in every eukaryotic cell) and might have also evolved direct sensitivity to stretch … Once its cortical position and mechanosensitivity were acquired, the actomyosin network could automatically fulfil an additional function: cell-shape maintenance — as any localized cell deformation would stretch the cortical actomyosin network and trigger an immediate compensatory contraction. This property would have arisen as a side-effect (a ‘spandrel’) of the presence of cortical actomyosin for membrane repair, and quickly proved advantageous.
An “anticipatory step”? “Pre-positioning”? Actomyosin “evolved” sensitivity to stretch? The position and mechanosensitivity “were acquired”? The network could “fulfil an additional function”? Sorry, but molecular machines (such as actomyosin) don’t “evolve” anything. There is more teleology packed into these few sentences than any medieval tract. And for good measure the authors also add the astonishing serendipity that this additional function “would have arisen as a side-effect.” That was lucky.
Once covering the cell cortex, the actomyosin network acquired the ability to deform the cell by localized contraction.
The actomyosin network “acquired the ability” to deform the cell by localized contraction? Smart move on the part of the network. But may we ask just how did that happen?
Based on the genomic study of the protist Naegleria which has a biphasic life cycle (alternating between an amoeboid and a flagellated phase), amoeboid locomotion has been proposed to be ancestral for eukaryotes. It might have evolved in confined interstitial environments, as it is particularly instrumental for cells which need to move through small, irregularly shaped spaces by exploratory deformation.
Amoeboid locomotion evolved “as it is particularly instrumental.” No infinitive form but this is no less teleological. Things don’t evolve because they are “instrumental.” What the authors fail to inform their readers of is that this would require an enormous number of random mutations.
One can hypothesize that, if stretch-sensitive calcium channels and cortical actomyosin were part of the ancestral eukaryotic molecular toolkit (as comparative genomics indicates), membrane deformation in a confined environment would probably trigger calcium influx by opening of stretch-sensitive channels, which would in turn induce broad actomyosin contraction across the deformed part of the cell cortex, global deformation and cell movement away from the source of pressure.
The concept of a “molecular toolkit” is standard in evolutionary thought, and another example teleological thinking.
One can thus propose that a simple ancestral form of amoeboid movement evolved as a natural consequence of the scenario outlined above for the origin of cortical actomyosin and the calcium — contraction coupling; once established, it could have been further elaborated.
Amoeboid movement evolved “as a natural consequence,” and “once established” was “further elaborated”? This is nothing more than teleological story-telling with no supporting evidence.
It is thus tempting to speculate that, once calcium signalling had gained control over primitive forms of amoeboid movement, the same signalling system started to modify ciliary beating, possibly for ‘switching’ between locomotor states.
Calcium signaling “gained control” and then “started to modify” ciliary beating “for ‘switching’ between locomotor states”? The “for switching” is yet another expression redolent of teleology, and “gained control” is an active move by the calcium signaling system. Pure, unadulterated, teleology.
Possibly, in ancestral eukaryotes calcium induced a relatively simple switch (such as ciliary arrest, as still seen in many animal cells and in Chlamydomonas in response to high Ca++ concentrations), which was then gradually modified into more subtle modulations of beating mode with a fast turnover of molecular actors mediated by differential addition, complementation and loss.
“Calcium induced a relatively simple switch”? Sorry, ions don’t induce switches, simple or otherwise. And the switch “was then gradually modified into more subtle modulations”? Note how the passive voice obviates those thorny details. The switch “was modified” conveniently omits the fact that such modification would have to occur via random mutation, one mutation at a time.
Alternatively, control of cilia by calcium could have evolved convergently — but such convergence would then have been remarkably ubiquitous, as there seems to be no eukaryotic flagellum that is not controlled by calcium in one way or another.
“Could have evolved convergently”? And exactly how would that happen? At least the authors then admit to the absurdity of that alternative.
Unfortunately, they lack such sensibility for the remainder of the paper. As we saw above, the paper is based on a sequence of teleological thinking. It falls into the evolutionary genre where evolution is taken, a priori, as a given. This going in assumption underwrites vast stretches of teleological thought, and cartoon-level story telling. Not only is there a lack of empirical support, but the genre is utterly unscientific, as revealed by even a mildly critical reading.
And needless to say, the paper does absolutely nothing to alleviate the problem we began with. The many leaps of logic and reasoning in the paper reveal all manner of monumental changes evolution requires to construct nerve cells and the action potential technology. We are not looking at a narrative of minute, gradual changes, each contributing to the overall fitness. Many, many simultaneous mutations are going to be needed. Even a conservative minimum number of 100 simultaneous mutations leads to the untenable result of a one in 10^800 chance of occurring.
It’s not going to happen.
Image credit: Nrets at en.wikipedia (Transferred from en.wikipedia) [CC BY 2.5], from Wikimedia Commons.
{kind=link}