 Evolution
Evolution
Kimberella — A Checkered History
Editor’s note: We are delighted to present a series of posts by paleontologist Günter Bechly on the Ediacaran organism Kimberella. If identified as an animal, it would “predate the Cambrian explosion of bilaterian animal phyla as a kind of ‘advance guard.’” The question is of interest for debates about evolution and arguments about intelligent design raised by Stephen Meyer, among others. For the first post in the series, see:
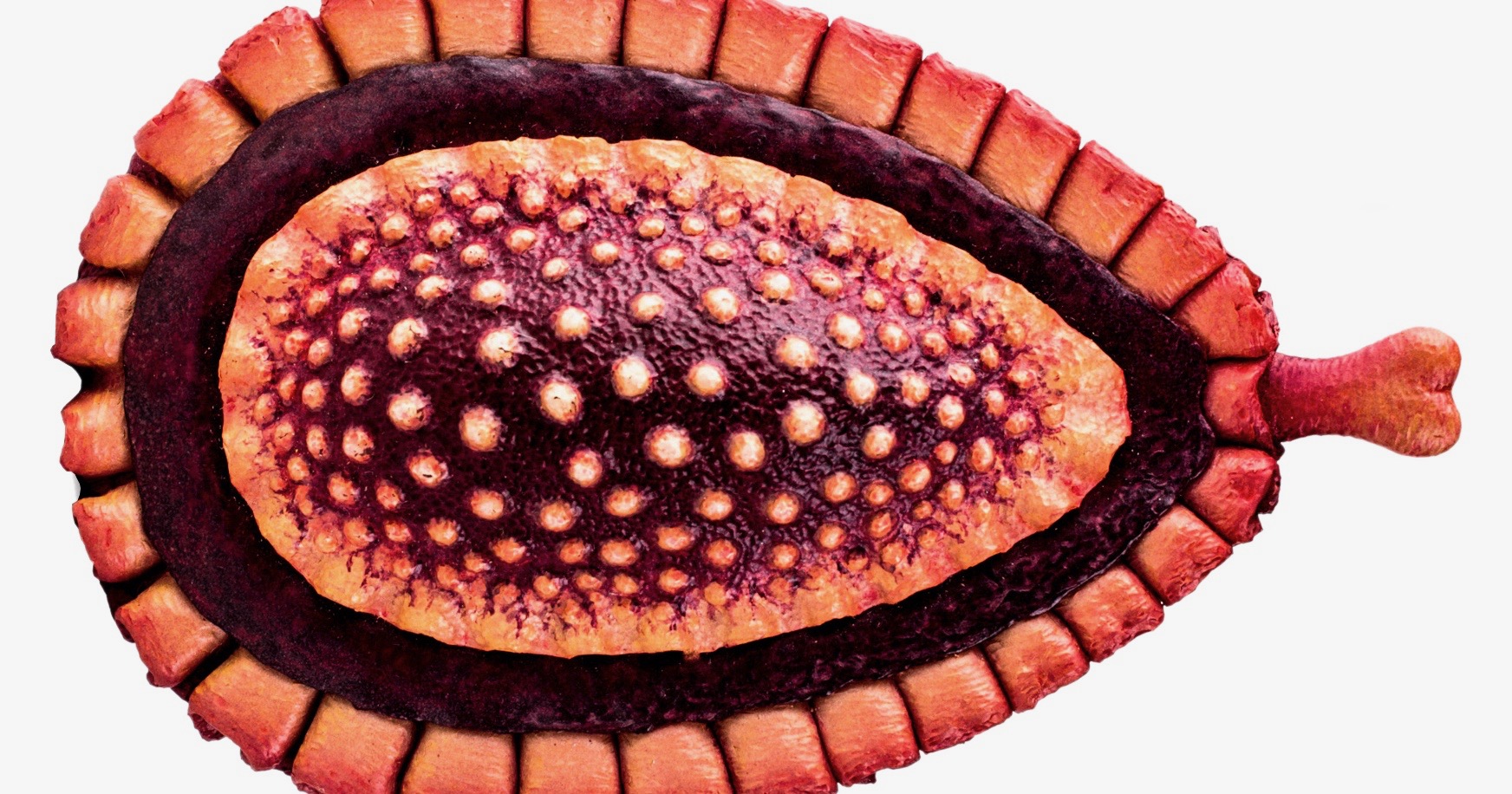
Kimberella is represented by relatively large ovoid fossils with complex structure from late Precambrian strata. Based on just a few relatively poorly preserved specimens from the Ediacara Hills of the Flinders Ranges in South Australia, Kimberella quadrata was first figured more than half a century ago by Glaessner & Daily (1959: pl. 47, fig. 9). The holotype specimen (no. SAM P12734) was described by Glaessner & Wade (1966) under the already occupied and thus invalid genus name Kimberia, in commemoration of John Kimber, who collected the first fossils of this organism and died tragically at age 38 during an expedition in South Australia in 1964. The taxon was re-described and renamed with the replacement genus name Kimberella by Wade (1972).
Radiometric Dating
These early works characterized Kimberella as a relatively large medusoid-like organisms related to modern jellyfish. The fossiliferous sandstones (Rawnsley Quartzite or Pound Quartzite) of the Ediacara Member are considered to be about 560-550 million years old. Unfortunately, though, due to a lack of suitable rocks there is no radiometric dating available yet.
In 1994 and subsequent years, Kimberella became much better known from more than thousand well-preserved specimens from the glauconitic mudstones and sandstones of the Zimnie Gory / Winter Coast (Zimnie Gory and Erga / Mezen Formations) and Solza River / Onega Peninsula (Verkhovka Formation) localities of the Arkhangelsk Region in the Russian White Sea area (Fedonkin & Waggoner 1997, Fedonkin et al. 2007b, Ivantsov et al. 2007, Ivantsov 2009). They are often found in association with Kimberichnus rasping traces and with the more enigmatic Ediacaran organisms like Dickinsonia and Tribrachidium.
Based on the radioisotope U-Pb age of zircon crystals from volcanic ash, these layers have been reliably dated to the Ediacaran epoch 4 (Upper Vendian) with an absolute age of 558 ± 1.0 to 555.3 ± 0.3 million years (Martin et al. 2000, Grazhdankin 2003, 2004). That is at least 6 million years older than the Nama assemblage in Namibia (549-543 mya). Based on this better-preserved White Sea material, Kimberella was re-interpreted as a benthic bilaterian mollusk-like animal (Fedonkin & Waggoner 1997, Waggoner 1998) that lived as a motile grazer. This result has been celebrated as a major breakthrough and as clear evidence for the presence of well-defined animal phyla in the Ediacaran (Seilacher 1999).
Kimberella fossils have also been reported from the Ediacaran Krol Formation from the Nainital District of Uttar Pradesh in India (Shanker & Mathur 1992, Shukla et al. 2006), but Fedonkin & Waggoner (1997) considered them to be questionable.
“Garden of Ediacara”
A new species, Kimberella persii, was recently described by Vaziri et al. (2018) based on nine poorly preserved specimens from the terminal Ediacaran of Kushk in the Chahmir basin of central Iran. If the attribution to Kimberella should be correct, then this would be the youngest record for this genus. However, this attribution is far from being well established, because all known specimens lack the tapered end and instead seem to have two equally broadened and rounded ends. Also, the segmented ridge along the midline and the lack of the lateral organic frill are quite different from the type species of Kimberella.
The maverick American paleontologist Mark McMenamin (2018), who coined the expression “Garden of Ediacara” and is well known for many original but highly controversial hypotheses, recently described relatively poorly preserved alleged specimens of Kimberella cf. quadrata from the Ediacaran Clemente Formation of the Sonoran desert in México. He also described from this locality Zirabagtaria ovata as a new form of solzid kimberellomorph, as well as Korifogrammia clementensis, which he considered as an intermediate between the annelid-like proarticulates (like Dickinsonia) and the mollusk-like Kimberella. McMenamin (2016) had already described an alleged aculiferan Clementechiton sonorensis McMenamin & Fleury from this Ediacaran locality, but his polyplacophoran reconstruction is highly dubious as it is based only on a single disarticulated body valve preserved as mold in sandstone, which in my view is much more likely just an over-interpreted ovoid imprint of uncertain origin.
Other Possible Relatives
Some other Ediacaran organism have been suggested as possible relatives of Kimberella. Fedonkin et al. (2007a: 276), Trusler et al. (2007), and Grazhdankin (2014) suggested that Brachina delicata Wade 1972 and Solza margarita Ivantsov et al. 2004 could be taphomorphs of Kimberella quadrata and thus represent synonyms of the same species, and Fedonkin also listed the problematic fossil Zolotytsia biserialis Fedonkin 1981 as a synonym. Grazhdankin (2014) noted that Parvancorina and Temnoxa, which were sometimes considered as proto-arthropods, show similarities to parts of Kimberella. Ivantsov (2010b, 2012) compared Solza margarita, Temnoxa molliuscula Ivantsov et al. 2004, and Armillifera parva Fedonkin 1980 with Kimberella, which were all sympatric contemporaries from the Vendian of Russia. However, he also found important anatomical differences, especially the glide symmetry in Armillifera, which he correctly considered as incompatible with a bilaterian mollusk bauplan. Nevertheless, Solza and Armillifera look strikingly similar in shape and tuberculated surface sculpture to smaller specimens of Kimberella preserved in dorsal aspect (Ivantsov 2010b: pl. 2).
A new higher taxon Kimberellomorpha was erected as unranked clade within Metazoa by Erwin et al. (2011) to accommodate Kimberella and the problematic Ediacaran fossil Solza margarita Ivantsov et al. 2004 (also see Fedonkin et al. 2007a: 286) from the Onega peninsula at the Russian White Sea region, which shares the ovoid body shape and tuberculate dorsal surface. Budd & Jensen (2017) casually remarked that the erection of a separate clade Kimberellomorpha was “perhaps not very helpful.”
Erwin et al. (2011) characterized these Kimberellomorpha as “centimeter-sized bilaterally symmetrical fossils with a crenulated margin interpreted as a frill surrounding a muscular foot, and a proboscis … commonly associated with radiating trace fossils that may represent feeding on microbial mats” and
oval-shaped, bilaterally symmetrical fossils composed of several morphologically distinct and concentrically-arranges zones. Outermost zone is typically smooth and crenulated, while innermost region is bordered by thin transverse wrinkles and contains a deep longitudinal invagination in the center. The anterior end is narrower and appears to house a retractable, arrow-shaped structure presumed to be responsible for the fan-shaped scratch marks (i.e., Radulichnus) sometimes associated with Kimberella.
Similarly, Laflamme et al. (2013) noted that “Kimberellomorpha … is a clade of oval-shaped bilaterally-symmetrical fossils with at least three concentrically arranged zones and clear anteroposterior differentiation …. A distal rasping radula-like appendage, combined with the close association of Kimberella with Radulichnus traces …, have resulted in this group being considered as stem group mollusks.”
Next, “Conflicting Views about Kimberella’s Ecology.”