 Evolution
Evolution
 Intelligent Design
Intelligent Design
#9 Story of 2021: Verdicts of “Poor Design” in Biology Have a Poor Track Record
Editor’s note: Welcome to an Evolution News tradition: a countdown of our Top 10 favorite stories of the past year, concluding on New Year’s Day. Our staff are enjoying the holidays, as we hope that you are, too! Help keep the daily voice of intelligent design going strong. Please give whatever you can to support the Center for Science & Culture before the end of the year!
The following was originally published on May 21, 2021.
In four previous posts (here, here, here, and here), I have commented on a TEDx talk by recent MIT bioengineering PhD Dr. Erika DeBenedictis, “It’s Time for Intelligent Design.” She makes a case that biology is poorly designed, and needs to have “intelligent design” added to it. But there is reason for caution in such an argument. I want to revisit three historical examples of so-called “poor design” in biology that turned out to not be so poorly designed. I revisit these to emphasize the need for caution in labeling biology “poorly designed” and to make the point that usually science just doesn’t have the whole story.
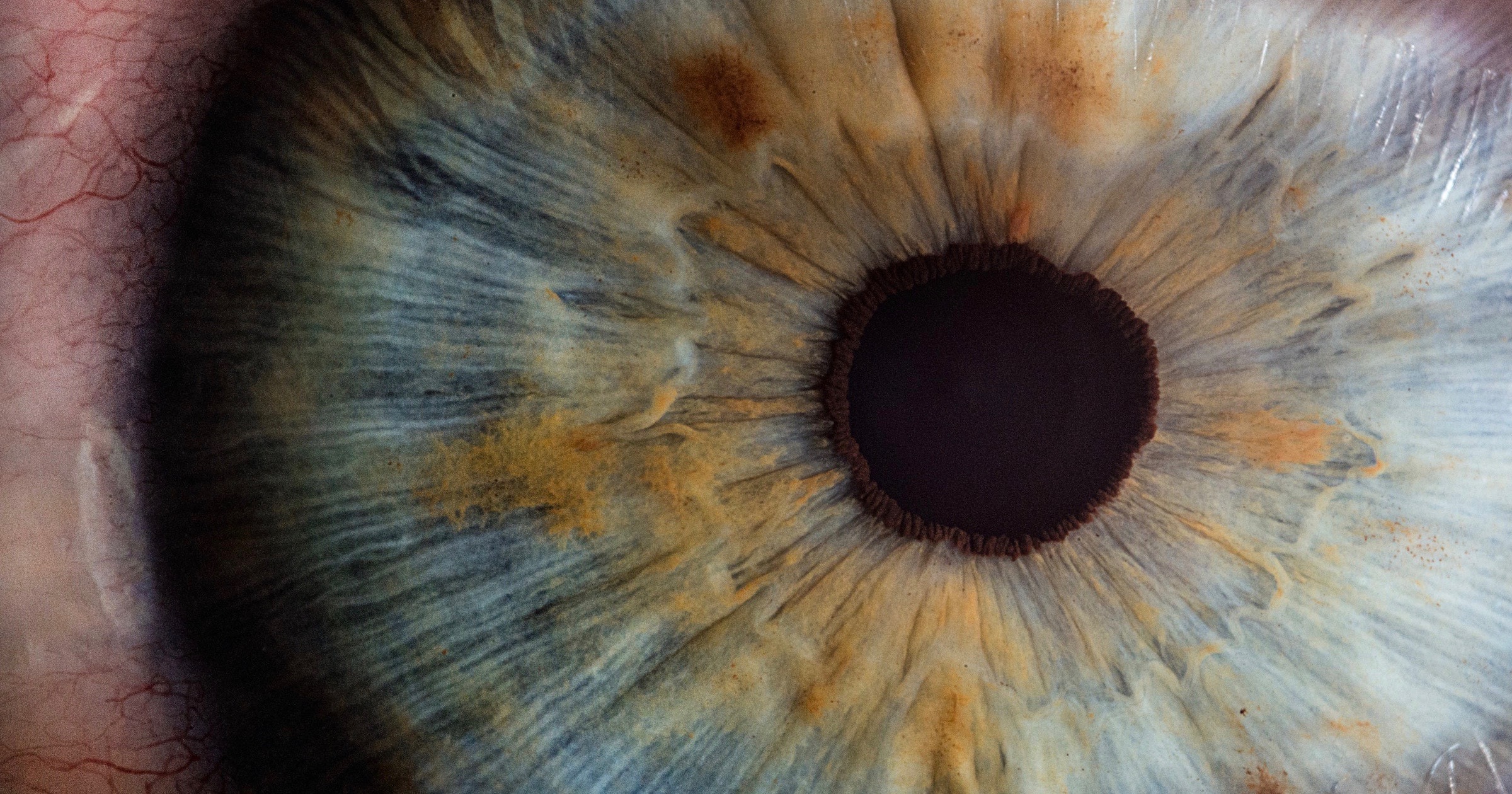
The Vertebrate Eye Wired Backwards
For years people cited the wiring of the vertebrate eye as evidence of “poor design” in biology. But starting in 2010 (Labin and Ribak) and continuing through 2014 researchers solved the mystery of the backwards wiring. “For the first time, we’ve explained why the retina is built backwards, with the neurons in front of the photoreceptors, rather than behind them. The retina is not just the simple detector and neural image processor, as believed until today,” said Erez Ribak, a professor at the Technion (Israel Institute of Technology). “Its optical structure is optimized for our vision purposes.” It turns out that the backwards wiring enables improvements in daytime vision without affecting night vision:
The findings presented here indicate that the spectral separation of light by Müller cells provides a mechanism to improve cone-mediated day vision, with minimal interference with rod-mediated night vision. This is achieved by wavelength sorting of incident light by the Müller cells. Light of relevant wavelengths for cone visual pigments is directed towards the cones, while light of wavelengths more suitable for rod vision is allowed to leak outside the Müller cells towards the surrounding rods. This is a novel mechanism that needs to be considered when visual phenomena concerning cone- and rod-mediated vision are analyzed.
(LABIN ET AL.)
The discovery of function for the optic nerve extending over the retina refuted the common objection that the vertebrate eye is “poorly designed.” It provided an excellent example of how this label often reflects an incomplete understanding of biology.
The Appendix Labeled as a Vestigial Organ
The misunderstood appendix, often vexed by the modern diet, has been labeled for many years by scientists and medical practitioners alike as an “evolutionary leftover.” Today, with an improved understanding of the microbiome and immune system, scientists now realize that the appendix is actually a safe house for normal gut bacteria. It reseeds the colon with normal gut bacteria following diarrheal washout and facilitates the development of the intestinal immune system. There you have it. Another example of the “poor design” label being incorrectly applied to an area of human physiology that was poorly understood. (Vitetta et al.; Laurin et al.; Gebbers and Laissue)
The GTP Proofreading Step as a Wasteful Side Reaction
The misguided verdict of “poor design” has extended even to core molecular machinery. Cells rely on extremely accurate protein translation to ensure the correct folding and function of proteins. The accuracy of protein translation depends upon a GTP hydrolysis mediated proofreading step that was once considered a “wasteful side reaction.” Uri Alon in his book An Introduction to Systems Biology explores how this reaction allows for a second discrimination step that results in multiplicative lowering of the error rate.
The fact that the modified tRNA can fall off seems wasteful because the correct tRNA can be lost. Moreover, to make c* costs energy: each amino acid incorporated into a protein requires hydrolysis of GTP which is about one ATP’s worth of energy. This cost adds up to a large part of the cell’s energy balance. However, it is precisely this design that generates high fidelity. The secret is that c* offers a second discrimination step: the wrong tRNA, once modified, can fall off the codon, but it cannot mount back on. (p. 121)
Assuming the GTP driven proofreading step was a wasteful side reaction is logical only if you think biology is the cobbled-together product of random Darwinian processes. In order to make better predictions, we must start with better assumptions.
In my last post, I explained why there is not sufficient knowledge of overlapping reading frames or adequate consideration of constraints to confidently state that INK4a and ARF are evidence of poor design. Historical examples also caution against prematurely labeling not-well-understood biological phenomena.
In the next post I will address how the assumption of overall good design provides a safe and rational justification for correcting corrupted biological information. I will agree with DeBenedictis that we shouldn’t wait around for natural selection and random mutation to fix instances of corruption. When the intended design has been damaged by random mutation, we can use intelligent design to restore its good intended design.