 Paleontology
Paleontology
Fossil Friday: Rhenocystis and the Controversial Calcichordate Hypothesis
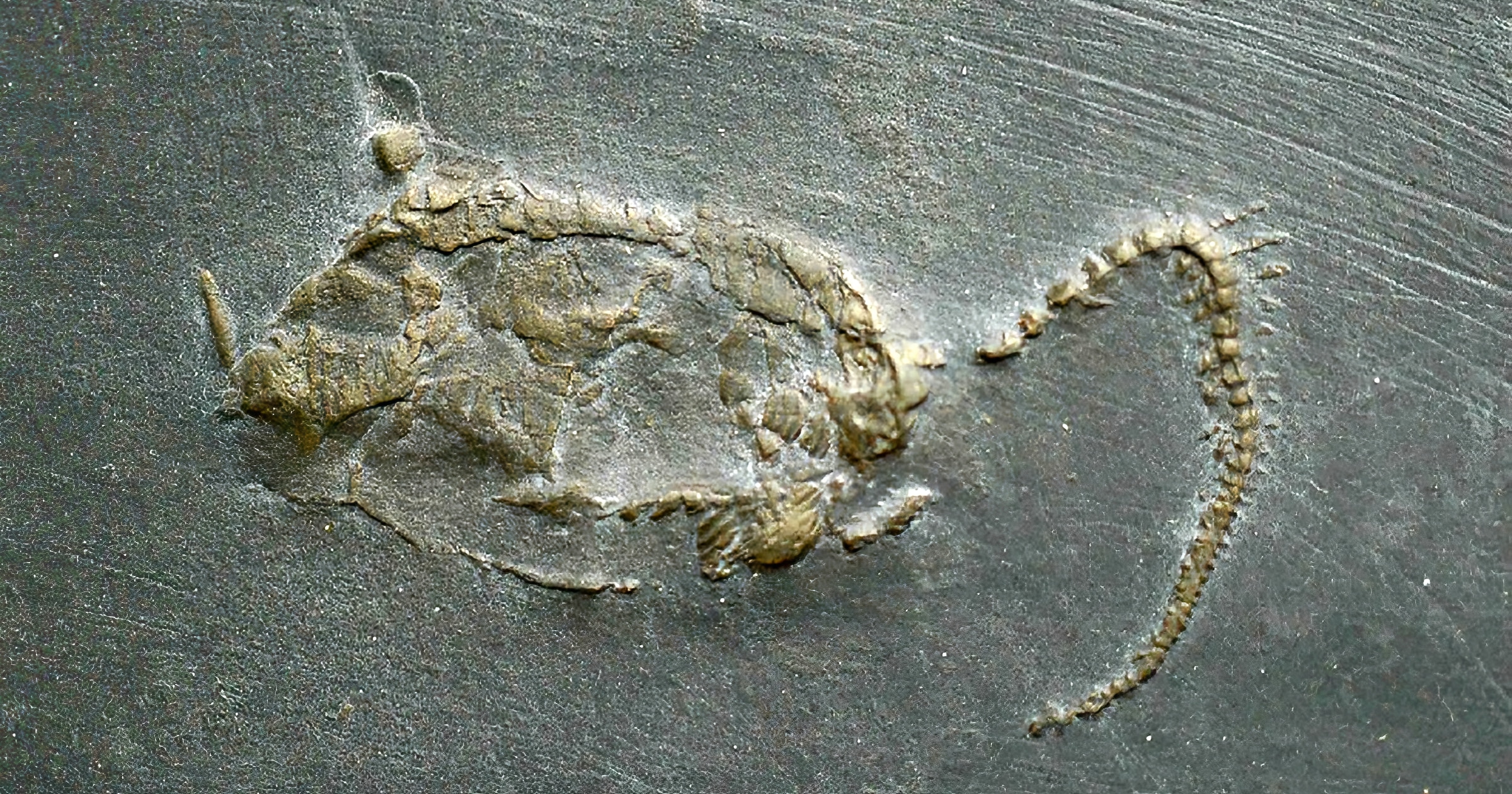
This Fossil Friday features the fossil echinoderm Rhenocystis latipendunculata from the 403 million-years-old Devonian Hunsrück shale in Germany. It looks a bit like a tadpole with body and tail, and this indeed points towards one of the great scientific controversies of the 20th century. The maverick paleontologist Richard Jefferies, who worked as a curator at the prestigious British Museum of Natural History, introduced and strongly promoted for 35 years a controversial theory for the origin of chordates, — the so-called ‘Calcichordate Hypothesis’ (Jefferies 1967, 1968, 1971, 1975, 1981, 1982, 1986, 1997, Jefferies et al. 1996, and Dominguez et al. 2002). He suggested that chordates are most closely related to echinoderms, and that lancelets, tunicates, and vertebrates independently evolved from a Paleozoic group of asymmetrical calcified organisms called carpoids (today named Homalozoa–Stylophora, including the subgroups Soluta, Cornuta, and Mitrata), which were usually considered as oldest and most basal echinoderms. Jefferies provided very elaborate reinterpretations of the anatomy of these fossils, in which he even claimed to recognize homologues of gill slits and the vertebrate brain with its main nerves. What he considered as a chordate tail, was considered by other paleontologists as an echinoderm feeding appendage. In other words: the expert disagreement was also about what is up and down and front and back in these fossil organisms, as well as the complete internal anatomy.
Hotly Debated
Jefferies’ calcichordate hypothesis was hotly debated in the late 1980s and 90s, but rejected by most other scientists as unfounded, debunked, and based on false methodology and phantastic speculation (Philip 1979, Fisher 1982, 1993, Jollie 1982, Sprinkle 1987, Peterson 1995, Nielsen 1995, Bergström et al. 1998, Ruta 1999, Simonetta et al. 1999, Lefebvre 2000, and Lefebvre et al. 2019). I still remember vividly that during my study at the University of Tübingen in the early 1990s, Jefferies’s hypothesis was just considered as an object of ridicule, together with other fringe ideas like Gardiner’s and Lovtrup’s haemothermia hypothesis of a close relationship of birds and mammals. The monumental textbook on the phylogenetic systematics of vertebrate animals authored by my greatly admired teacher Gerhard Mickoleit (2004), did not even consider Jefferies’ hypothesis worth mentioning. Critically referring to Jefferies’ kind of reasoning, Peterson (1995) boldly concluded that “fossils are unable to dictate relationships”. Few other paleontologists supported Jefferies’ interpretations. A notable exception were Eaton (1970a, 1970b), Shu et al. (2001), and especially Nature editor in chief Henry Gee, who strongly defended the calcichordate hypothesis in a long chapter of his book (Gee 1996). He later suggested a kind of compromise hypothesis (Gee 2001). In a study co-authored with Jefferies and published in Nature, Dominguez et al. (2002) described a stylophoran fossil as clearly possessing the predicted paired gill slits and attributed it to the stem group of tunicates. This latest and potentially final defense of the calcichordate theory was criticized as highly controversial by Bertrand Lefebvre (quoted by MacAndrew 2003), who rather considered the alleged gill slits as part of the genital organs (gonoducts).
An Interesting Twist
An interesting twist to this whole story was based on the circumstance that the opponents of the calcichordate hypothesis considered as their most substantive critique that this hypothesis implies a closest relationship of tunicates with vertebrates, which was generally considered as refuted by numerous independent phylogenetic studies (Peterson 1995, Donoghue et al. 2003). However, the comprehensive phylogenomic analysis by Delsuc et al. (2006) and several other modern studies (e.g., Bourlat et al. 2006, Putnam et al. 2008, Sawalla & Smith 2008, Giribet 2018, and Kocot et al. 2018; but see Stach 2008, Zupanc 2008, and Braun et al. 2020 for an opposing view) surprisingly suggested that tunicates are indeed closer related to vertebrates. Delsuc et al. (2006) explicitly mention that this would be compatible with Jefferies’ theory. However, they also emphasized that this does not mean that Jefferies’ interpretation is necessarily true. Most paleontologists and evolutionary biologists still considered stylophorans as echinoderms and the calcichordate hypothesis as a failed and obsolete idea. This was mainly for the following two reasons (Lefebvre et al. 2019): Even though the calcichordate hypothesis correctly predicted a sister group relationship of tunicates and vertebrates, it also relies on a sister group relationship of echinodermates and chordates that was refuted by modern phylogenomics, which rather suggests that echinodermates are the sister group to hemichordates. Furthermore, the unique stereom skeleton of echinodernates would have to be interpreted as secondarily reduced in chordates, which is an unparsimonious ad hoc assumption, for which there is no independent evidence. Finally, Lefebvre et al. (2019) could document the typical water-canals in the appendage, which strongly suggests a homology with an echinoderm arm and not with a chordate tail. It looks like we can definitely put the calcichordate hypothesis to rest. Even though the early critics seemingly were proven right, this does of course not excuse their intolerant opposition to a unconventional new view in a time when the jury was still out.
What is our take home message from this controversy? Well, at least these points: Fossils are often open to very different alternative reconstructions and interpretations (Donoghue & Purnell 2009), and phylogenetic trees often change with every new data instead of converging to the one true tree of life. This suggests that we should not place too much confidence in the currently favored scientific consensus in these fields. Also, unconventional ideas are often met with fierce opposition and ridicule in the scientific community, which is something we ID proponents know all too well. Maybe both points are related: if the evidence is weaker than pretended, the subconscious insecurity may lead to a more angry defense of the preferred but relatively weakly founded position. Keep this in mind when discussing with an angry defender of Darwinism next time.
References
- Bergström J, Naumann WW, Viehweg J & Martí-Mus M 1998. Conodonts, calcichordates and the origin of vertebrates. Mitteilungen aus dem Museum für Naturkunde in Berlin (Geowissenschaftliche Reihe) 1(1), 81–92. DOI: https://doi.org/10.1002/mmng.19980010106
- Bourlat SJ, Juliusdottir T, Lowe CJ et al. 2006. Deuterostome phylogeny reveals monophyletic chordates and the new phylum Xenoturbellida. Nature 444, 85–88. DOI: https://doi.org/10.1038/nature05241
- Braun K, Leubner F & Stach T 2020. Phylogenetic analysis of phenotypic characters of Tunicata supports basal Appendicularia and monophyletic Ascidiacea. Cladistics 36(3), 259–300. DOI: https://doi.org/10.1111/cla.12405
- Delsuc F, Brinkmann H, Chourrout D & Philippe H 2006. Tunicates and not cephalochordates are the closest living relatives of vertebrates. Nature 439(7079), 965–968. DOI: https://doi.org/10.1038/nature04336
- Dominguez P, Jacobson AG & Jefferies RPS 2002. Paired gill slits in a fossil with a calcite skeleton. Nature 417, 841–844. DOI: https://doi.org/10.1038/nature00805
- Donoghue PCJ, Smith MP & Sansom IJ 2003. The origin and early evolution of chordates: molecular clocks and the fossil record. pp. 190–223 in: Donoghue PCJ & Smith MP (eds). Telling the evolutionary time: molecular clocks and the fossil record. Systematics Association Special Volume Series 66. CRC Press, Boca Raton (FL), viii+296 pp.
- Donoghue PCJ & Purnell MA 2009. Distinguishing heat from light in debate over controversial fossils. Bioessays 31(2), 178–189. DOI: https://doi.org/10.1002/bies.200800128
- Eaton TH 1970a. [Book review] The Subphylum Calcichordata (Jefferies, 1967), Primitive Fossil Chordates with Echinoderm Affinities by R.P.S. Jefferies. Journal of Paleontology 44(1), 168. https://www.jstor.org/stable/1302511
- Eaton TH 1970b. The stem-tail problem and the ancestry of chordates. Journal of Palaeontology 44(5), 969–979. https://www.jstor.org/stable/1302733
- Fisher DC 1982. Stylophoran skeletal cristallography: testing the calcichordate theory of vertebrate origins. Abstracts with Programs of the Geological Society of America 14, 488.
- Fisher DC 1993. Life orientation of mitrate stylophorans and its implications for the calcichordate theory of vertebrate origins. Abstracts with Programs of the Geological Society of America 25, A-105.
- Gee H 1996. Jefferies’ Calcichordate Theory. pp. 201-286 in: Gee H. Before the Backbone: Views on the origin of the vertebrates. Chapman & Hall, London (UK), xx+346 pp.
- Gee H 2001. Deuterostome phylogeny: the context for the origin and evolution of chordates. pp. 1–14 in: Ahlberg PE (ed.). Major Events in Early Vertebrate Evolution: Palaeontology, Phylogeny, Genetics and Development. Systematics Association Special Volume Series 61. Taylor & Francis, London (UK) & New York (NY), 418 pp.
- Giribet G 2018. Phylogenomics resolves the evolutionary chronicle of our squirting closest relatives. BMC Biology 16:49, 1–3. DOI: https://doi.org/10.1186/s12915-018-0517-4
- Jefferies RPS 1967. Some fossil chordates with echinoderm affinities. Symposia of the Zoological Society of London 20, 163–208.
- Jefferies RPS 1968. The subphylum Calcichordata (Jefferies 1967) primitive fossil chordates with echinoderm affinities. Bulletin of the British Museum (Natural History) Geology 16(6), 243–339. DOI: https://doi.org/10.5962/p.313838
- Jefferies RPS 1971. Some Comments on the Origin of Chordates. Journal of Paleontology 45(5), 910–912. https://www.jstor.org/stable/1302789
- Jefferies RPS 1975. Fossil evidence concerning the origin of the chordates. Symposia of the Zoological Society of London 36, 253–318.
- Jefferies RPS 1981. In defence of the calcichordates. Zoological Journal of the Linnean Society 73(4): 351–396. DOI: https://doi.org/10.1111/j.1096-3642.1981.tb01601.x
- Jefferies RPS 1982. The calcichordate controversy – comments on Notocarpos garratti Philip. Alcheringa 6, 78. DOI: https://doi.org/10.1080/03115518208565422
- Jefferies RPS 1986. The Ancestry of the Vertebrates. Cambridge University Press & British Museum, Cambridge & London (UK), 376 pp.
- Jefferies RPS 1997. A defence of the calcichordates. Lethaia 30(1), 1–10. DOI: https://doi.org/10.1111/j.1502-3931.1997.tb00437.x
- Jefferies RPS, Brown NA & Daley PE 1996. The Early Phylogeny of Chordates and Echinoderms and the Origin of Chordate Left-Right Asymmetry and Bilateral Symmetry. Acta Zoologica 77(2), 101–122. DOI: https://doi.org/10.1111/j.1463-6395.1996.tb01256.x
- Jollie M 1982. What are the ‘Calcichordata’? and the larger question of the origin of Chordates. Zoological Journal of the Linnean Society 75(2), 167–188. DOI: https://doi.org/10.1111/j.1096-3642.1982.tb01946.x
- Kocot KM, Tassia MG, Halanych KM & Swalla BJ 2018. Phylogenomics offers resolution of major tunicate relationships. Molecular Phylogenetics and Evolution 121, 166–173. DOI: https://doi.org/10.1016/j.ympev.2018.01.005
- Lefebvre B 2000. Homologies in Stylophora: A test of the ‘Calcichordate Theory’. Geobios 33(3) 359–364. DOI: https://doi.org/10.1016/S0016-6995(00)80163-1
- Lefebvre B, Guensburg TE, Martin ELO, Mooi R, Nardin E, Nohejlová M, Saleh F, Kouraïss K, El Hariri K & David B 2019. Exceptionally preserved soft parts in fossils from the Lower Ordovician of Morocco clarify stylophoran affinities within basal deuterostomes. Geobios 52, 27–36. DOI: https://doi.org/10.1016/j.geobios.2018.11.001
- MacAndrew A 2003. A very early chordate fossil. Evolution Pages. http://www.evolutionpages.com/Jaekelocarpus%20oklahomensis.htm
- Mickoleit G 2004. Phylogenetische Systematik der Wirbeltiere. Verlag Dr. Friedrich Pfeil, Munich (Germany), 671 pp. https://pfeil-verlag.de/publikationen/phylogenetische-systematik-der-wirbeltiere/
- Nielsen C 1995. Animal Evolution. Oxford University Press, Oxford (UK), 467 pp. [Third Edition 2012]
- Peterson KJ 1995. A phylogenetic test of the calcichordate scenario. Lethaia 28(1), 25–38. DOI: https://doi.org/10.1111/j.1502-3931.1995.tb01591.x
- Philip GM 1979. Carpoids — Echinoderms or Chordates? Biological Reviews 54(4), 439–471. DOI: https://doi.org/10.1111/j.1469-185X.1979.tb00845.x
- Putnam NH, Butts T, Ferrier DEK et al. 2008. The amphioxus genome and the evolution of the chordate karyotype. Nature 453, 1064–1071. DOI: https://doi.org/10.1038/nature06967
- Ruta M 1999. Brief review of the stylophoran debate. Evolution & Development 1(2), 123–135. DOI: https://doi.org/10.1046/j.1525-142x.1999.99008.x
- Swalla BJ & Smith AB 2008. Deciphering deuterostome phylogeny: molecular, morphological and palaeontological perspectives. Philosophical Transactions of the Royal Society B 363, 1557–1568. DOI: https://doi.org/10.1098/rstb.2007.2246
- Shu D-G, Conway Morris S, Han J, Chen L, Zhang X-L, Zhang Z-F, Liu H-Q, Li Y & Liu J-N 2001. Primitive deuterostomes from the Chengjiang Lagerstätte (Lower Cambrian, China). Nature 414(6862), 419–424. DOI: https://doi.org/10.1038/35106514
- Simonetta AM, Pucci A & Dzik J 1999. Hypotheses on the origin and early evolution of chordates in the light of recent zoological and palaeontological evidence. Italian Journal of Zoology 66(2), 99–119. DOI: https://doi.org/10.1080/11250009909356245
- Sprinkle J 1987. The Calcichordate Theory. Science 236(4807), 1476. DOI: https://doi.org/10.1126/science.236.4807.1476 [DOI-link broken, therefore use https://pubmed.ncbi.nlm.nih.gov/17793239/]
- Stach T 2008. Chordate phylogeny and evolution: a not so simple three-taxon problem. Journal of Zoology 276(2), 117–141. DOI: https://doi.org/10.1111/j.1469-7998.2008.00497.x
- Zupanc GKH 2008. Chordate evolution: not just a molecular phylogenetics problem. Journal of Zoology 276(2), 115–116. DOI: https://doi.org/10.1111/j.1469-7998.2008.00513.x