 Paleontology
Paleontology
Fossil Friday: Fossil Hyraxes and the Abrupt Origin of Hyracoidea
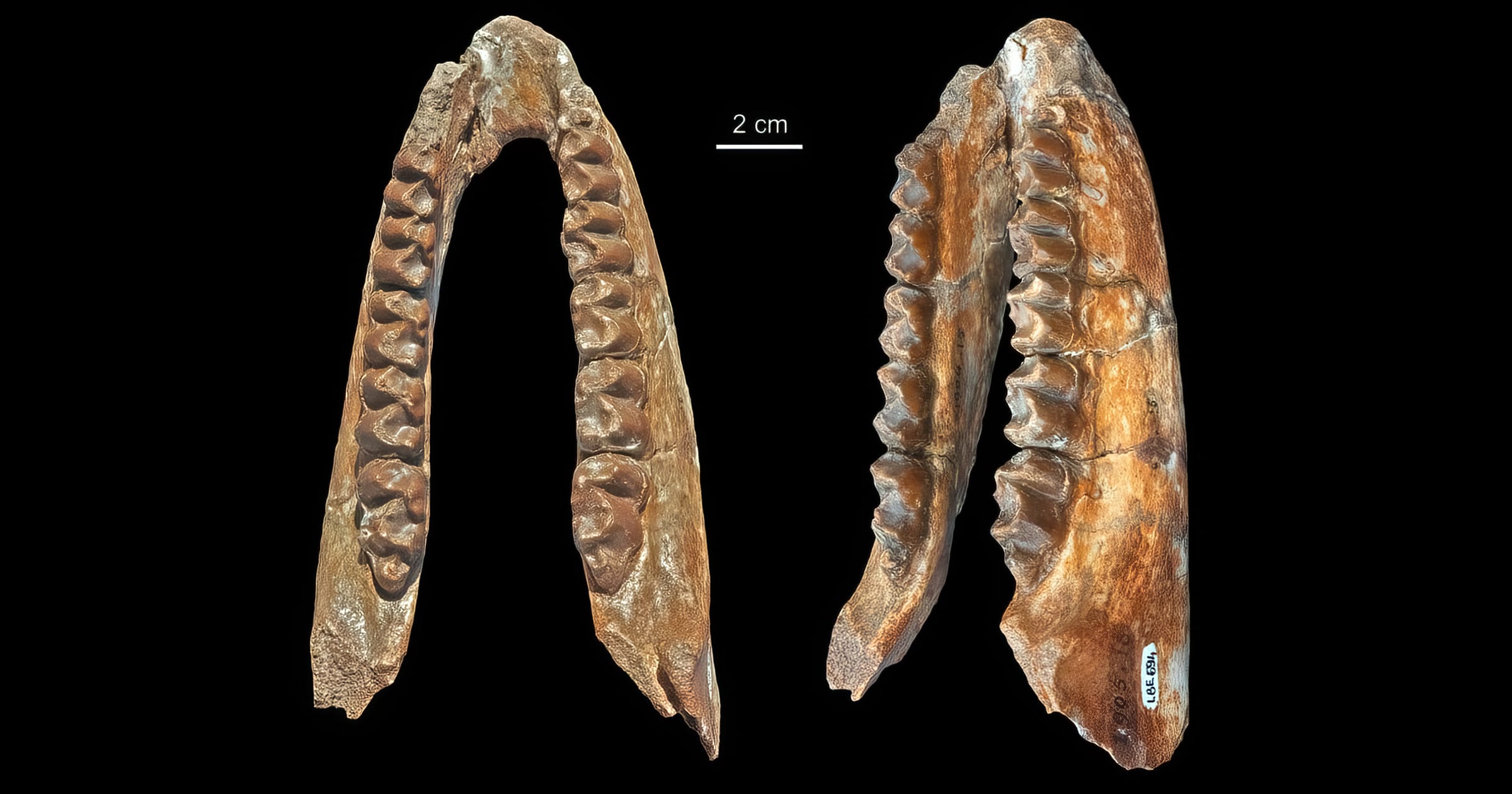
This Fossil Friday features the giant hyrax Titanohyrax andrewsi from the Early Oligocene of Fayum in Egypt (Tabuce 2016), because today we look into the origins of the placental mammal order Hyracoidea. This order only includes the herbivorous Afro-Arabian hyraxes that look rather like marmots even though they are believed to be close relatives of elephants and manatees. Whereas the five living species of the single surviving family Procaviidae look very similar, fossil hyraxes especially in the Paleogene were much more diverse with five extinct families (Geniohyiidae, Namahyracidae, Pliohyracidae, Sagatheriidae, and Titanohyracidae), sometimes subsumed in a single paraphyletic family Pliohyracidae. These included forms from the size of a mouse to that of a rhino (Tabuce 2016), which occupied very different ecological niches (Rasmussen & Simons 2000). The strange genus Rukwalorax described by Stevens et al. (2009), based on a single tooth from the Late Oligocene of Tanzania, could represent another extinct family and the oldest small-bodied hyracoid from East Africa. The Paleogene fossil record of hyraxes is surprisingly rich and diverse (Rasmussen 1989, Fischer 1992: table 1, Tabuce et al. 2008, Barrow et al. 2010: fig. 16, Rasmussen & Gutiérrez 2010).
The possibly oldest fossil record of Hyracoidea is represented by an isolated molar tooth of Seggeurius spec. from the earliest Eocene of the Ouled-Abdoun basin in Morocco (Gheerbrant et al. 2003, Asher & Seiffert 2010: fig. 46.2, Seiffert 2010a), which is dated to 55.8 million years. The second oldest find is Seggeurius amourensis from the middle Ypresian (ca. 52 mya) El Kohol Formation of the southern Atlas in Algeria (Mahboubi et al. 1986, Court & Mahboubi 1993, Seiffert 2010a, Benoit et al. 2016). Some of the other oldest fossils of Hyracoidea were also found in the Early Eocene of Algeria at Gour Lazib (Sudre 1979, Court & Mahboubi 1993, Adaci et al. 2007, Tabuce et al. 2001, 2011, Seiffert 2010a), which have been dated to a late Ypresian / early Lutetian age (ca. 52-46 mya) and include the species Megalohyrax gevini, Microhyrax lavocati, and Titanohyrax mongereaui. Titanohyrax tantulus from the Early Eocene of Chambi in Tunisia (Hartenberger et al. 1985, Court & Hartenberger 1992, Hartenberger et al. 2001) is of about the same age (Barrow et al. 2010, Seiffert 2010a). Numerous other very old hyraxes have been found in Late Eocene (Early Priabonian) layers of the Fayum Depression in Egypt (Barrow et al. 2010), such as the 37 million-year-old Dimaitherium, and the Late Eocene (Lutetian and Bartonian) of Sperrgebiet in Namibia (Pickford et al. 2008, Pickford 2015).
Abrupt and Diverse
We can conclude that hyraxes appeared abruptly and with a surprising diversity about 56 million years ago in the very window of time when most other orders of placental mammals appeared for the first time as well. There are no fossils that show an assumed gradual development of hyraxes from Late Cretaceous stem eutherians via stem afrotherians etc. The diversity was most developed very early in the Middle to Late Eocene and is comparatively small today, which is about the opposite of what should be expected in a Darwinian scenario. Tabuce et al. (2011) therefore admitted: “To conclude, the diversity of hyracoids in the first part of the Maghrebian Eocene is remarkable and surprising at such an early age.” Words like “surprising” are code in the technical evolutionary literature for facts that disagree with Darwinian expectations and predictions, to avoid clearly stating the embarrassing fact of the matter.
Hyraxes were initially wrongly believed to be related to rodents. Since George Cuvier’s (1884: 120) time until relatively recently, hyraxes have often been considered to be more closely related to the odd-toed ungulates (Perissodactyla) in a group called Altungulata or Pantomesaxonia (e.g., Fischer 1986, 1992, Fischer & Tassy 1993, Prothero & Schoch 1989, McKenna & Bell 1997, Halliday et al. 2015), while other researchers instead have followed George Gaylord Simpson (1945) in considering them as close relatives of manatees and elephants in a group called Paenungulata (Sale 1960). Studies disagreed about the precise position of hyraxes, with most studies suggesting a basal position, while others suggested a closer relationship with either elephants (Sale 1960) or more rarely with sea cows (Seiffert 2010b, Benoit et al. 2016). Modern phylogenomic studies confirmed the monophyly of Paenungulata and placed them in the Afrotheria clade of African mammals (Asher et al. 2003, Nishihara et al. 2005, Seiffert 2003, 2007, Asher & Seiffert 2010, O’Leary et al. 2013, Cooper et al. 2014, Heritage et al. 2020). Among the very few anatomical features that might support Afrotheria (Tabuce et al. 2007, 2008) is the increased number of thoracolumbar vertebrae (Sánchez-Villagra et al. 2007) and the lack of a scrotum, but the latter similarity is somewhat incongruent as it is absent in the aardvarks, which are supposed to be nested within afrotherians. Recently, genetic evidence has been found that indeed suggests that the reduction of the testicular descent happened independently within Afrotheria (Sharma et al. 2018).
Not Based on Common Ancestry
Once again, anatomical similarity turns out not to be based on common ancestry. This is supported even more by the striking fact that there are three groups of mammals that independently produced a hyracoid-like morphology, so that they were initially misidentified as hyraxes:
- The Eocene early Hippomorpha (“horses”) like Hyracotherium.
- The fossil elephant shrew family Miohyracidae (see Bechly 2022).
- The family Archaeohyracidae of the South American ungulate clade Notungulata. A recent study by Avilla & Mothé (2021) suggested that notungulates are indeed related to afrotherian hyracoids, but this result was immediately disputed by Kramarz & MacPhee (2022), who found them nested within the unrelated Boreoeutheria instead. See why I got personally frustrated with phylogenetics as a wannabe science?
Of course, it is only we “nitpicking” intelligent design proponents who point out such incongruences, while Darwinists generally see no problem at all. The theory must be correct, therefore any conflicting evidence must be wrong and explained away, following the Procrustean solution of shoehorning the data until they fit.
Next Fossil Friday we will look into the early fossil history of another member of the Afrotheria, the order Sirenia, which includes manatees and dugongs.
References
- Adaci M, Tabuce R, Mebrouk F, Bensalah M, Fabre P-H, Hautier L, Jaeger J-J, Lazzari V, Mahboubi M, Marivaux L, Otero O, Peigné S & Tong H 2007. New Palaeogene vertebrate sites from the Gour Lazib area (northwestern Sahara, Algeria). Comptes Rendus Palevol6(8), 535–544. DOI: https://doi.org/10.1016/j.crpv.2007.09.001
- Asher RJ & Seiffert ER 2010. Systematics of Endemic African Mammals. Chapter 46, pp. 911–928 in: Werdelin L & Sanders WJ (eds). Cenozoic Mammals of Africa. University of California Press, Berkeley (CA), 1008 pp. https://doi.org/10.1525/california/9780520257214.003.0046
- Asher RJ, Novacek MJ & Geisher JH 2003. Relationships of Endemic African Mammals and Their Fossil Relatives Based on Morphological and Molecular Evidence. Journal of Mammalian Evolution 10(1/2), 131–194. DOI: https://doi.org/10.1023/A:1025504124129
- Avilla LS & Mothé D 2021. Out of Africa: A New Afrotheria Lineage Rises From Extinct South American Mammals. Frontiers in Ecology and Evolution 9:654302, 1–14. https://doi.org/10.3389/fevo.2021.654302
- Barrow E, Seiffert ER & Simons EL 2010. A primitive hyracoid (Mammalia, Paenungulata) from the early Priabonian (Late Eocene) of Egypt. Journal of Systematic Palaeontology8(2), 213–244. DOI: https://doi.org/10.1080/14772010903450407
- Bechly G 2022. Fossil Friday: Fossil Elephant Shrews and the Abrupt Origin of Macroscelidea. Evolution News December 30, 2022. https://evolutionnews.org/2022/12/fossil-friday-fossil-elephant-shrews-and-the-abrupt-origin-of-macroscelidea/
- Benoit J, Crochet J-Y, Mahboubi M, Jaeger J-J, Bensalah M, Adaci M & Tabuce R 2015. New material of Seggeurius amourensis (Paenungulata, Hyracoidea), including a partial skull with intact basicranium. Journal of Vertebrate Paleontology 36(1):e1034358, 1–17. DOI: https://doi.org/10.1080/02724634.2015.1034358
- Cooper LN, Seiffert ER, Clementz M, Madar SI, Bajpai S, Hussain ST & Thewissen JGM 2014. Anthracobunids from the Middle Eocene of India and Pakistan are stem perissodactyls. PLoS ONE 9(10):e109232, 1–15. DOI: https://doi.org/10.1371/journal.pone.0109232
- Court N & Hartenberger J-L 1992. A new species of the hyracoid mammal Titanohyrax from the Eocene of Tunisia. Palaeontology 35(2), 309–317. https://www.palass.org/publications/palaeontology-journal/archive/35/2/article_pp309-317
- Court N & Mahboubi M 1993. Reassessment of Lower Eocene Seggeurius amourensis: Aspects of Primitive Dental Morphology in the Mammalian Order Hyracoidea. Journal of Paleontology 67(5), 889–893. http://www.jstor.org/stable/1306052
- Cuvier Baron 1884. The Animal Kingdom, arranged after its organization. New edition. Allen & Co, London (UK).
- Fischer MS 1986. Die Stellung der Schliefer (Hyracoidea) im phylogenetischen System der Eutheria. Courier Forschungsinstitut Senckenberg 84, 1–132.
- Fischer MS 1992. Hyracoidea. Handbuch der Zoologie, Band VIII, Teilband 58. De Gruyter, Berlin (DE), 184 pp. DOI: https://doi.org/10.1515/9783110879711
- Fischer MS & Tassy P 1993. The interrelation between Proboscidea, Sirenia, Hyracoidea, and Mesaxonia: the morphological evidence. pp. 217–234 in: Szalay FS, Novacek MJ & McKenna MC (eds). Mammal Phylogeny, Vol. 2: Placentals. Springer, New York (NY), 332 pp.
- Gheerbrant E, Sudre J, Cappetta H, Mourer-Chauviré C, Bourdon E, Iarochene M, Amaghzaz M & Bouya B 2003. Les localités à mammifères des carrières de Grand Daoui, bassin des Ouled Abdoun, Maroc, Yprésien : premier état des lieux. Bulletin de la Societe Geologique de France 174(3), 279–293. https://www.researchgate.net/publication/27598819
- Halliday TJD, Upchurch P & Goswami A 2015. Resolving the relationships of Paleocene placental mammals. Biological Reviews 92(1), 521–550. DOI: https://doi.org/10.1111/brv.12242
- Hartenberger J-L, Martinez C & Ben Saïd A 1985. Découverte de mammifères d’âge Éocène inférieur en Tunisie centrale. Comptes Rendus de l’Académie des Sciences Paris Série 2 301, 649–652. https://gallica.bnf.fr/ark:/12148/bpt6k6372204t/f665.image
- Hartenberger J-L, Crochet J-Y, Martinez C, Marandat B & Sige B 2001. The Eocene mammalian fauna of Chambi (Tunisia) in its geological context. pp. 237–250 in: Gunnell GF (ed.). Eocene Biodiversity: Unusual Occurrences and Rarely Sampled Habitats. Springer, New York (NY), xxi+442 pp. DOI: https://doi.org/10.1007/978-1-4615-1271-4_9
- Heritage S, Rayaleh H, Awaleh DG & Rathbun GB 2020. New records of a lost species and a geographic range expansion for sengis in the Horn of Africa. PeerJ 8: e9652, 1–38. DOI: https://doi.org/10.7717/peerj.9652
- Kramarz AG & MacPhee RDE 2022. Did some extinct South American native ungulates arise from an afrothere ancestor? A critical appraisal of Avilla and Mothé’s (2021) Sudamericungulata – Panameridiungulata hypothesis. Journal of Mammalian Evolutionpreprint 2022. https://doi.org/10.1007/s10914-022-09633-5
- Mahboubi M, Ameur R, Crochet JM & Jaeger JJ 1986. El Kohol (Saharan Atlas, Algeria): A New Eocene Mammal Locality in Northwestern Africa. Palaeontographica Abt. A 192, 15–49. https://www.schweizerbart.de/papers/pala/detail/A192/71186/
- McKenna MC & Bell SK 1997. Classification of Mammals Above the Species Level. Colombia University Press, New York, 631 pp.
- Nishihara H, Satta Y, Nikaido M, Thewissen JG, Stanhope MJ & Okada N 2005. A retroposon analysis of Afrotherian phylogeny. Molecular Biology and Evolution 22(9), 1823–1833. DOI: https://doi.org/10.1093/molbev/msi179
- O’Leary MA, Bloch JI, Flynn JJ et al. 2013. The Placental Mammal Ancestor and the Post–K-Pg Radiation of Placentals. Science 339 (6120), 662–667. DOI: https://doi.org/10.1126/science.1229237
- Pickford M 2015. New Titanohyracidae (Hyracoidea: Afrotheria) from the Late Eocene of Namibia. Communications of the Geological Survey of Namibia 16, 200–214. https://www.mme.gov.na/files/publications/6d9_Comms%20GSN%2016,%202015%20Pickford%20Hyracoidea,200-214.pdf
- Pickford M, Senut B, Morales J, Mein P & Sánchez IM 2008. Mammals from the Lutetian of Namibia. Memoirs of the Geological Survey of Namibia 20, 465–514. https://www.mme.gov.na/files/publications/a93_Memoir%2020_2008_Pickford&Senut_Geology%20and%20Palaeobiology%20of%20the%20Northern%20Sperrgebiet.pdf
- Prothero DR & Schoch RM 1989. Origin and Evolution of the Perissodactyla: Summary and Synthesis. pp. 504–537 in Prothero DR & Schoch RM (eds.). The Evolution of Perissodactyls. Oxford University Press, New York (NY), 560 pp. http://www.donaldprothero.com/files/47440280.pdf
- Rasmussen DT 1989. The evolution of the Hyracoidea: a review of the fossil evidence. pp. 57–76 in Prothero DR & Schoch RM (eds.). The Evolution of Perissodactyls. Oxford University Press, New York (NY), 560 pp.
- Rasmussen DT & Simons EL 2000. Ecomorphological diversity among Paleogene hyracoids (Mammalia): a new cursorial browser from the Fayum, Egypt. Journal of Vertebrate Paleontology 20(1), 167–176. https://www.jstor.org/stable/4524072
- Rasmussen DT & Gutiérrez M 2010. Hyracoidea. Chapter 13, pp. 123–145 in: in: Werdelin L & Sanders WJ (eds). Cenozoic Mammals of Africa. University of California Press, Berkeley (CA), 1008 pp. DOI: https://doi.org/10.1525/california/9780520257214.003.0013
- Sale JB 1960. The Hyracoidea: A Review of the Systematic Position and Biology of the Hyrax. Journal of the East Africa Natural History Society 23(5):102, 185–188. https://www.biodiversitylibrary.org/part/140662
- Sánchez‐Villagra MR, Narita Y & Kuratani S 2007. Thoracolumbar vertebral number: The first skeletal synapomorphy for afrotherian mammals. Systematics and Biodiversity 5(1), 1–7. DOI: https://doi.org/10.1017/s1477200006002258
- Seiffert ER 2003. A phylogenetic analysis of living and extinct afrotherian mammals. Unpublished PhD dissertation, Duke University.
- Seiffert ER 2007. A new estimate of afrotherian phylogeny based on simultaneous analysis of genomic, morphological, and fossil evidence. BMC Evolutionary Biology 7(1):224, 1–13. DOI: https://doi.org/10.1186/1471-2148-7-224
- Seiffert ER 2010a. Chronology of paleogene mammal localities. Chapter 2, pp. 18–26 in: Werdelin L & Sanders WJ (eds). Cenozoic Mammals of Africa. University of California Press, Berkeley (CA), 1008 pp. DOI: https://doi.org/10.1525/california/9780520257214.003.0002
- Seiffert ER 2010b. The oldest and youngest records of afrosoricid placentals from the Fayum Depression of northern Egypt. Acta Palaeontologica Polonica 55(4), 599–616. DOI: https://doi.org/10.4202/app.2010.0023
- Sharma V, Lehmann T, Stuckas H, Funke L & Hiller M 2018. Loss of RXFP2 and INSL3genes in Afrotheria shows that testicular descent is the ancestral condition in placental mammals. PLoS Biology 16(6):e2005293, 1–22. DOI: https://doi.org/10.1371/journal.pbio.2005293
- Simpson GG 1945. The Principles of Classification and a Classification of the Mammals. Bulletin American Museum of Natural History 85, ix+350 pp. http://hdl.handle.net/2246/1104
- Stevens NJ, O’Connor PM, Roberts EM & Gottfried MD 2009. A hyracoid from the Late Oligocene Red Sandstone Group of Tanzania, Rukwalorax jinokitana (gen. and sp. nov.). Journal of Vertebrate Paleontology 29(3), 972–975. DOI: https://doi.org/10.1671/039.029.0302
- Sudre J 1979. Nouveaux Mammiferes Eocenes du Sahara Occidental. Palaeovertebrata9(3), 83–115. https://www.palaeovertebrata.com/Articles/sendFile/76/published_article
- Tabuce R 2016. A mandible of the hyracoid mammal Titanohyrax andrewsi in the collections of the Muséum National d’Histoire Naturelle, Paris (France) with a reassessment of the species. Palaeovertebrata 40(1):e4, 1–12. DOI: https://doi.org/10.18563/pv.40.1.e4
- Tabuce R, Mahboubi M & Sudre J 2001. Reassessment of the Algerian Eocene Hyracoid Microhyrax. Consequences on the early diversity and basal phylogeny of the Order Hyracoidea (Mammalia). Eclogae Geologicae Helvetiae 94(3), 537–545. DOI: http://doi.org/10.5169/seals-168910
- Tabuce R, Marivaux L, Adaci M, Bensalah M, Hartenberger JL, Mahboubi M, Mebrouk F, Tafforeau P & Jaeger J-J 2007. Early Tertiary mammals from north Africa reinforce the molecular Afrotheria clade. Proceedings of the Royal Society of London B 274(1614), 1159–1166. DOI: https://doi.org/10.1098/rspb.2006.0229
- Tabuce R, Asher RJ & Lehmann T 2008. Afrotherian mammals: a review of current data. Mammalia 72(1), 2–14. DOI: https://doi.org/10.1515/MAMM.2008.004
- Tabuce R, Charruault A-L, Adaci M, Bensalah M, Ali MBH, Essid EM, Marivaux L, Vianey-Liaud M & Mahboubi M 2011. The early Eocene radiation of Hyracoidea (Mammalia, Afrotheria): New fieldwork evidence from northwestern Africa. pp. 161–162 in: The World at the Time of Messel: Puzzles in Palaeobiology, Palaeoenvironment and the History of Early Primates. 22nd International Senckenberg Conference, Frankfurt a.M. (DE), Abstract Volume. https://www.researchgate.net/publication/271604890, https://www.academia.edu/24508589/