 Evolution
Evolution
 Intelligent Design
Intelligent Design
Peer-Reviewed Paper Shows Vertebrate Embryonic Variation Contradicts Common Ancestry
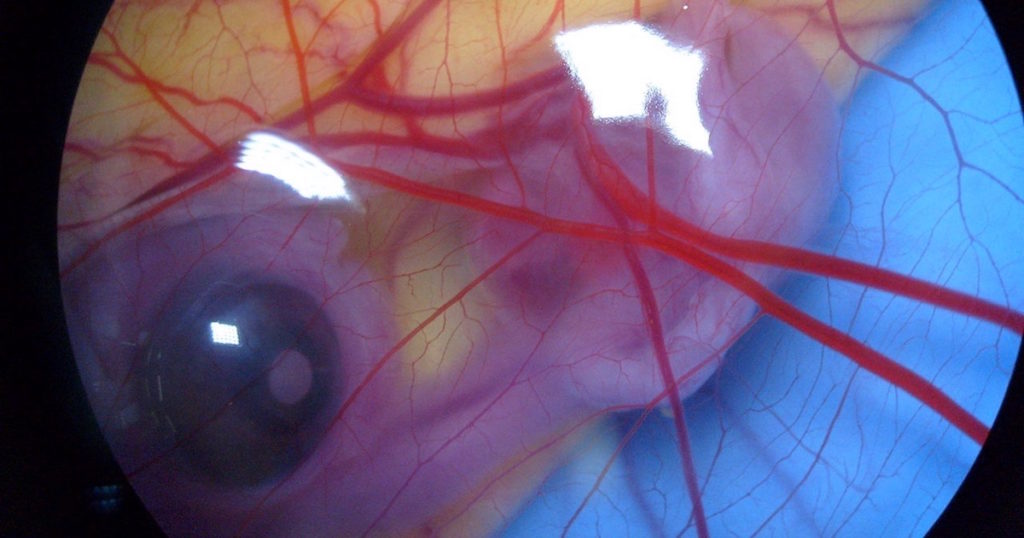
Evolutionary biologists often argue that vertebrate embryos develop in highly similar manners, reflecting their common ancestry. But a peer-reviewed paper in BIO-Complexity, by David Swift (author of Evolution Under the Microscope), provides an insightful review on the subject. The article is titled, “The Diverse Early Embryonic Development of Vertebrates and Implications Regarding Their Ancestry.” Swift shows that, despite common claims, vertebrates do not develop similarly, according to the predictions of evolution.
He opens by framing his thesis:
It is well known that the embryonic development of vertebrates from different classes (e.g., fish, reptiles, mammals) pass through a “phylotypic stage” when they look similar, and this apparent homology is widely seen as evidence of their common ancestry. However, despite their morphological similarities, and contrary to evolutionary expectations, the phylotypic stages of different vertebrate classes arise in radically diverse ways. This diversity clearly counters the superficial appearance of homology of the phylotypic stage, and the plain inference is that vertebrates have not evolved from a common vertebrate ancestor. The diversity extends through all stages of early development — including cleavage and formation of the blastula, gastrulation, neurulation, and formation of the gut and extraembryonic membranes.
Now intelligent design does not require common ancestry to be false, but even a guided form of common ancestry might lead to different predictions from strictly unguided descent with modification. Thus, Darwin and subsequent evolutionists such as Ernst Haeckel found embryology to be a crucial line of evidence supporting Darwin’s thesis. According to Swift, common ancestry predicts that vertebrate development should exhibit striking homologies. But more than that, “If common ancestry is the explanation for homologies, not only should homologous organs be derived from equivalent embryonic tissues (the cardinal criterion for homology) but they should also develop by comparable processes.”
Very Different Pathways
The crux of Swift’s thesis is that although vertebrate embryos do pass through a similar “phylotypic stage,” the pathways of development are very different:
[D]espite their morphological similarities and contrary to evolutionary expectations, the striking fact is that the “phylotypic stages” of different groups of vertebrates arise in remarkably diverse ways, even with key tissues such as the germ layers (see below) deriving from completely different early embryonic sources. These observations clearly refute the presumed evolutionary homology of the vertebrate phylotypic stage, and hence undermine the inference of common ancestry based on that supposed homology. [Emphasis in the original.]
He reprints the “hourglass model” of vertebrate development and points out that this is merely an “observation” about development — not an explanation of how it arose:
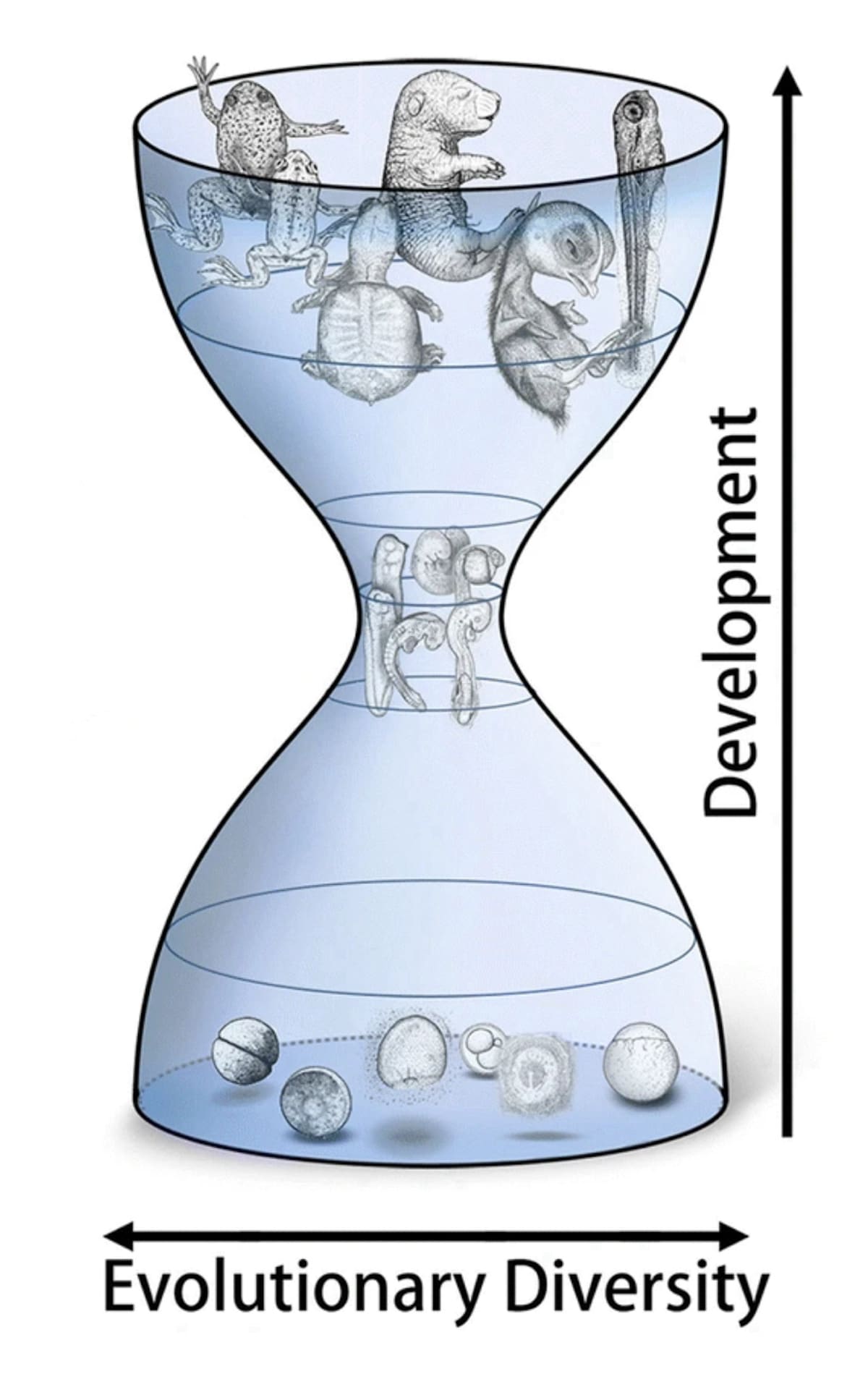
One of the key stages of development that leads to this similar “phylotypic stage” is gastrulation, which Swift notes is crucial because it establishes the basic body plan and “leads to the establishment of the germ layers — ectoderm, mesoderm and endoderm — from which all of the body’s tissues are derived.”
Differences in Vertebrate Development
Swift cites various specific differences in vertebrate development to make his case, especially in gastrulation. He predicts that “from an evolutionary perspective we would surely expect gastrulation to be ‘conserved’” but finds that “for almost all of the major classes of vertebrates” there are key differences in gastrulation, including “the mechanism of gastrulation is significantly different from any of the others,” and “the source tissues of the germ layers are different.”
After reviewing mechanisms of gastrulation in various vertebrate classes, Swift notes that “the wide variety of structures of the blastulas of different classes of vertebrate challenges the view that the resultant embryonic tissues can be considered equivalent or homologous.” Specifically, in different types of vertebrates different parts of the blastula ultimately become the embryo itself. He describes these differences as follows:
- Chondrichthyans (lancelets): “It is a one-cell thick epithelial layer, forming the upper surface of the blastula.”
- Teleosts (bony fish, e.g., zebrafish): “It is a multiple-cell layer, beneath the overlying enveloping layer.”
- Amphibians: “It is the whole of the blastula, comprising the multilayered dome of the upper hemisphere and the mass of cells in the lower hemisphere.”
- Reptiles and birds: “It is the upper surface of the blastula, comprising a single-cell thick epithelial layer, overlying the hypoblast.”
- Placental mammals (e.g., primates): “It is part of the inner cell mass, within the outer trophoblast.”
He cites further differences between which cells become the endoderm and which become the mesoderm, noting:
- in amniotes (reptiles, birds, mammals) cells that are internalized arise from a central area of the epiblast, i.e., the presumptive endoderm and mesoderm are surrounded by presumptive ectoderm; whereas
- in anamniotes (chondrichthyans, teleosts, amphibians) the cells that internalize are from the edge of the epiblast, i.e., the presumptive endoderm and mesoderm surround the presumptive ectoderm.
Swift summarizes major differences in the mechanisms of gastrulation as follows:
- Chondrichthyans: by cells rolling over a posterior overhang of the epiblast.
- Teleosts: by involution around the edges of the epiblast as it spreads around the yolk.
- Amphibians: by involution through an annular blastopore.
- Reptiles: by involution through a canal-like blastopore.
- Birds: by cells ingressing through a primitive streak, formation of the primitive streak being accompanied by growth of an underlying endoblast.
- Placental mammals: by cells ingressing through a primitive streak.
“Three Substantial Distinctions”
Swift thus finds that these diverse modes of development cannot be considered homologous:
In the light of these three substantial distinctions — the different overall structure of the blastulas, the different parts of the blastula that become the embryo, and the different relative positions of the presumptive ectoderm and mesoderm/endoderm in amniotes and anamniotes — there is no doubt that the tissues that become the embryo are not equivalent, and hence are far from being homologous across the various vertebrate classes.
According to Swift, these fundamental differences in early vertebrate mechanisms of development during gastrulation suggest that vertebrates do not share a common ancestry:
The straightforward conclusion to draw from this radical diversity of their early embryonic development is that it shows the vertebrates have not evolved from a common vertebrate ancestor. This conclusion can be avoided only if there are credible explanations for how such diversity of early development might have arisen from the development prevailing in a common ancestor (whether or not similar to present-day cephalochordates) in an evolutionary way, via changes that (i) had a realistic probability of occurring, (ii) maintained viability, and (iii) offered, in most cases, significant advantage that could be favored by natural selection.
Meeting these evolutionary requirements poses a great challenge, however. Swift quotes a prominent developmental biologist, Rudolf Raff, who wrote: “One might reasonably expect mechanisms of early development to be especially resistant to modification because all subsequent development derives from early processes.” Swift calls this a “commonsense conclusion” because the complexity of vertebrate development demands that many coordinated modifications would be required to fundamentally change how development proceeds. He thus finds that “because of the interdependence of the mechanisms that are involved, constructive changes to embryonic development must entail coordinated production of and/or changes to several genes, e.g., for transcription factors and the DNA sequences on which they act, which is prohibitively improbable.”
This leads to a “waiting time” problem where multiple coordinated change would be required to transition from one developmental regime to another, posing “a formidable challenge to supposed evolutionary scenarios” as generating these changes would be “generally far in excess of the time available.”