 Evolution
Evolution
 News Media
News Media
Derbyshire VI: Behe’s Bacterial Flagellum — Still Stirring Up Trouble for Darwin’s Defenders
John Derbyshire is on The Corner arguing that we can never safely infer that certain biological structures were designed. To a reader who asserted that organizational complexity cannot arise from impersonal processes, Derbyshire replies, “How do you know it can’t? It is true that the genesis of organizational complexity is not currently well understood; but to leap from that to telling me we shall NEVER be able to find a natural-law explanation for it is just dogma.”
Derbyshire’s argument is worth confronting because it represents the opinion of leading Darwinists. Biologist Kenneth Miller, for instance, routinely makes just such an argument. Design theorist William Dembski responds thus:
Miller claims that the problem with anti-evolutionists like Michael Behe and me is a failure of imagination — that we personally cannot “imagine how evolutionary mechanisms might have produced a certain species, organ, or structure.” He then emphasizes that such claims are “personal,” merely pointing up the limitations of those who make them. Let’s get real. The problem is not that we in the intelligent design community, whom Miller incorrectly calls “anti-evolutionists,” just can’t imagine how those systems arose.
The problem is that Ken Miller and the entire biological community haven’t figured out how those systems arose. It’s not a question of personal incredulity but of global disciplinary failure (the discipline here being biology) and gross theoretical inadequacy (the theory here being Darwin’s).
The particular mechanism Miller has in view here is the bacterial flagellum. Click here and scroll down for a good, brief description and animation of the bacterial flagellum, and here for an enlarged view with its parts labeled. Biochemist Michael Behe made this little engine that could famous by showing that it was irreducibly complex, like a mouse trap: “If any one of the components of the mousetrap (the base, hammer, spring, catch, or holding bar) is removed, then the trap does not function.” With even four of these parts, it’s utterly useless. The mousetrap is irreducibly complex.
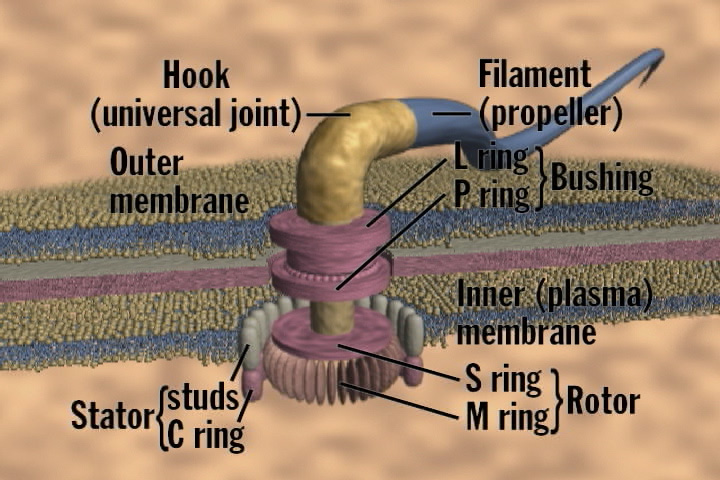{kind=link}
What does irreducible complexity have to do with Darwinian evolution? Evolution by mutation and natural selection must proceed by one slight, functional improvement at a time. So how can it build an irreducibly complex propeller motor one step at a time if the motor can’t propel at all until all of its parts are in place? It can’t. Something else built it.
Behe’s argument doesn’t assume that none of the other parts could ever be used for anything else. The spring on a mousetrap could be taken and used in some other device. The base with cheese on it could feed a mouse. Several but not all of the parts of a bacterial flagellum — while completely useless as a rotary propulsion machine — can be used to transport proteins across a membrane. But this hardly provides a credible Darwinian pathway.
Imagine if a boy told a girl he could climb to Mars because there supposedly existed a natural ladder stretching from one planet to the other? The girl is skeptical, pointing out that nobody on earth has ever found such a ladder. The boy screams, “That’s an argument from ignorance! Scientists are finding all sorts of new things all the time. Look! The moon! The moon is one step along the way. You see, everything is falling into place.” The Darwinists’ desperate efforts to spin away the clear significance of the bacterial flagellum is strangely akin to this sort of reasoning.
Darwin’s theory, without which nothing in biology is supposed to make sense, in fact offers no insight into how the flagellum arose. If the biological community had even an inkling of how such systems arose by naturalistic mechanisms, Miller would not — a full six years after the publication of Darwin’s Black Box by Michael Behe — be lamely gesturing at the type three secretory system as a possible evolutionary precursor to the flagellum.
Miller and Derbyshire are like the boy convinced of the natural ladder to Mars, who finds the moon and yells “Ah ha! Now who dares to play the skeptic!” Well, design theorists do. Consider this passage from a peer-edited paper by biologist Scott Minnich and philosopher of science Stephen Meyer, in which they discuss recent evidence for the delicately orchestrated and information-rich proteins of the bacterial flagellum:
[I]f anything, TTSSs [Type Three Secretory Systems] generate more complications than solutions to this question. As shown here, possessing multiple TTSSs causes interference. If not segregated one or both systems are lost. Additionally, the other thirty proteins in the flagellar motor (that are not present in the TTSS) are unique to the motor and are not found in any other living system. From whence, then were these protein parts co-opted?
Also, even if all the protein parts were somehow available to make a flagellar motor during the evolution of life, the parts would need to be assembled in the correct temporal sequence similar to the way an automobile is assembled in factory. Yet, to choreograph the assembly of the parts of the flagellar motor, present-day bacteria need an elaborate system of genetic instructions as well as many other protein machines to time the expression of those assembly instructions.
Arguably, this system is itself irreducibly complex. In any case, the co-option argument tacitly presupposes the need for the very thing it seeks to explain–a functionally interdependent system of proteins.
Now one letter writer, responding to Behe’s column in The New York Times, complained that we only describe the bacterial flagellum as an outboard motor because we have no better analogy, not because it is an outboard motor. The letter writer argues that believing this molecular motor was designed is like the astronomer Percival Lowell mistaking Martian canyons for canals. The suggestion is that false design inferences have been made, so surely all of the design inferences in the natural sciences are false. In this case, the fallacious argument — a favorite among Darwinists — is doubly silly because the Martian canyons turned out to be far simpler, far less specified, than engineered canals; while the bacterial flagellum has turned out to be far more sophisticated than our outboard motors.
As Minnich and Meyer note, the discovery of molecular motors is opening a whole new field, where biology and engineering meet:
To paraphrase the original rendition of the Department of Energy’s Genomes to Life web site, “the molecular machines present in the simplest cells, produced by evolution, dwarf the engineering feats of the 20th century.” The dissection of the complexity and sophistication of … machines like the bacterial flagellum are indeed a testimony to the power of modern molecular biological techniques. Yet, the elegant structural properties, efficiency, and the highly controlled genetic programming to produce these machines was neither anticipated nor predicted. The potential applications of this knowledge are legion and have spawned a new discipline focused on nanotechnology.
One needn’t go far for examples. Here at Physics Today, well trained physicists are standing around this astonishing little machine, the bacterial flagellum, like neighborhood mechanics getting a chance to take apart and learn from a NASCAR racing engine.
Derbyshire, Miller and other Darwinists are mixed up about the direction of things. The more we KNOW about the bacterial flagellum, the less and less it is anything the Darwninian mechanism could produce. Moreover, there are strongly affirmative grounds for inferring design from the presence of irreducibly complex machines and circuits. Every time we know the causal history of an irreducibly complex system (like the NASCAR racing engine or an electronic circuit), it always turn out to have been the product of an intelligent cause.
Finally, the list of known biological mechanisms that appear irreducibly complex isn’t shrinking, it’s growing.