 Evolution
Evolution
 Faith & Science
Faith & Science
 Life Sciences
Life Sciences
Beginning to Decipher the SINE Signal
Remember the analogy of the two moons I used yesterday to discuss the distribution of SINEs in the mouse and rat genomes? Well, I am going to use it again today, but only for a moment.
Moon Mysteries and the Lunarlogos Foundation
Suppose you are keenly interested in the topography of one of the moons, named Y6-9. Suppose also that the books you first select to read on the topic are popular works, written by “experts” who are “living legends.” As you read through the works, you find paragraphs here and there about how utterly decrepit Y6-9 is, and how this space body exemplifies eons of random events. The authors argue that we already knew all there was to know about that moon back in 1859, and that the evidence demonstrates either that God doesn’t exist or that the deity left the cosmos to itself after the Big Bang.
You find, however, that these books almost totally ignore the findings of the billion-dollar missions sent to the surface of Y6-9 since the 1960s. Indeed, there is next to nothing in them about Y6-9’s actual geology.
So you contact the Lunarlogos Foundation, a Christian group that promotes such books. You tell them that you have a few specific questions about the Y6-9 mission findings. The response you get is that because you are a layman, you would not be able to comprehend the details. Besides, the Lunarlogos folks say, the mainstream experts have spoken authoritatively about the subject and that should be enough for you. As a consolation, though, they send you a CD that has songs that are sung by one of their founding members.
Somewhat disgruntled, you decide to spend a day at a university library. You ask a librarian for maps of Y6-9 and technical journals that discuss its features. An hour or so later, with stacks of data before you, something catches your eye–something never mentioned in any of the books you’ve read. Sitting in a Y6-9 crater is a large monolith. High resolution photos reveal it to be rectangular in shape, with a polished surface, and composed of some dense black material. This must be a mistake, you think. So you look at other craters on Y6-9 and many of them also contain the same kind of monolith. You discern their overall distribution to be non-random–and the monoliths themselves are highly non-random. Then, after consulting the literature, you learn: The existence of such objects has been known for over two decades. In fact, one of the experts of Lunarlogos wrote about them in the technical reports of the Y6-9 probe missions.
ou decide to write about what you have learned, citing the relevant literature in case someone might want to read about this topic themselves. After posting what you write on the Internet, Lunarlogos posts their reply. Their response reads something like this:
Okay. Sure. There are obnoxious monoliths littering Y6-9…everybody knows this. In fact, there are about a million of them. But they got there because of degenerative cosmic processes. While many of the structures Mr. X mentioned are suggestive of some possibly unknown cause that we have never denied, it is almost certain that much, if not most, of the Y6-9 surface is without any remarkable features. Besides, why would God put them there? They are simply nonsensical.
We have one more thing to say. We don’t appreciate how disrespectful Mr. X has been to our team of experts. Although Mr. X is a Ph.D. planetary scientist, he is not as qualified to write on this subject as scientists approved by Lunarlogos. So we ask him, for the sake of having meaningful dialogue: Please stop writing about this subject.
A Lunarlogos sympathizer writes on another blog:
We think you’re a nice guy, but your arguments are insane.
What would you think?
Then someone unaffiliated with Lunarlogos brings something to your attention. He shows you a map of the sister moon of Y6-9, called Q7-10. You are aware that Y6-9 and Q7-10 went their separate ways after a cosmic collision twenty-two million years ago. But something strange catches your eye. Q7-10 has polished black monoliths, too, except that they are pyramids instead of rectangles. That’s not the weirdest thing, though. The weirdest thing is that the geographical distribution of the monoliths on Y6-9 very nearly matches the geographical distribution of the monoliths on Q7-10.
Now what would you think?
That Strange SINE Signal…Again
The almost one-to-one correspondence of mouse-specific and rat-specific SINE insertion events along homologous regions of the two genomes is almost as remarkable as the matching geographical distributions of the monoliths in the analogy of the two moons. Remember the graph (from Figure 9c of Ref. 1):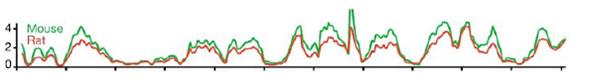
We have two genomes that went their separate ways 22 million years ago. We have two lineages that have been subjected to different historical events. Yet, when we compare the chromosome locations of mouse B1s/B2s/B4s with those of rat IDs, they look almost the same. Where the ID SINEs rise in density, so do the B1s/B2s/B4s SINEs; where the ID SINE levels decrease, so also do the B1s/B2s/B4s SINE levels. Independent mutational events have generated equivalent genomic patterns. How can we causally account for this striking pattern?
In the paper written by Francis Collins and his colleagues, under the heading Co-localization of SINEs in rat and mouse1, we read:
“The cause of the unusual distribution patterns of SINEs…is apparently a conserved feature, independent of the primary sequence of the SINE…” (Italics mine.)
Let’s unpack this part of the sentence. We have:
1) A cause of some sort.
2) A cause that is conserved between the mouse and rat.
3) A cause that is independent of SINE primary DNA sequences.
That’s all very well and good, but the specific cause is never mentioned. Where, then, can we find it?
Thinking Like a Darwinian
Experts such as John Avise, Francisco Ayala, Francis Collins, and Darrel Falk tell us that we must think like Darwinians before we can begin to make sense of the data, since nothing else is scientific, or indeed even reasonable. So let’s play along and think like Darwinians, limiting ourselves to what Collins and his colleagues have authoritatively provided. Recall that they are:
- Chance mutations continually degrade genomes that are largely junk
- SINEs are for the most part nonsensical junk
- Natural selection is the sole creative force in evolution
- Except when genetic drift (neutral evolution) is also a factor
We can call this conceptual scheme the “BioLogos box.”
We’ll start, then, with chance mutations. We know that the enzymes encoded by the L1 retrotransposon copy and paste SINEs into mammalian genomes. So perhaps this is the causative agent that acts independently of primary DNA sequence? And since L1 is present in all mammalian genomes, we may just be on the trail of the “conserved cause.”
But wait. L1 also mobilizes itself. This is a problem, for when we compare LINE and SINE distributions along chromosomes, it is clear that in the regions where the former is abundant the latter is not, and vice versa. Remember the graph (from Figure 9d of Ref. 1):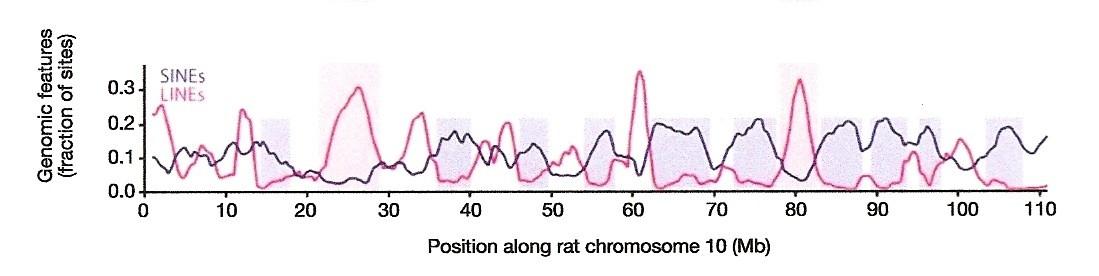
But we have no plausible mechanistic explanation for why the mouse L1 machinery would have pasted B1s/B2s/B4s–over twenty-two million years, no less–into the same general locations and at much the same densities, as the rat L1 machinery pasted ID elements over the same period of time.
Not to fear. We still have to consider that worker of miracles, natural selection. This mechanism eliminates harmful features while preserving those that enhance survival. So let’s construct a hypothesis: Mouse and rat SINE distributions reflect the differential removal of these DNA repeats from regions where their presence would be harmful. In other words, we predict that sequences where mouse B1s/B2s/B4s and rat IDs peak in density are segments of the genome that are largely junk; conversely, in the sections where these SINEs taper off, functional coding regions are to be found.
Does this hypothesis point in the right causal direction? I don’t think so. Here is why. Remember the statement made by Falk in defense of Ayala contra Meyer:
He [Ayala] does say that on average there are about 40 copies of Alu sequences between every two genes, but this is simply a fact.
Well, both Falk and Ayala are correct–and that is the problem with the selection hypothesis. Protein-coding genes make up only ~1.5% of the mammalian genome. Where do the peaks of B1s/B2s/B4s and IDs occur along the mouse and rat chromosomes, respectively? In and around the ~1.5% of the genome that is protein-coding. Remember the following statement in the sentence of the Nature paper quoted above1:
The cause of the unusual distribution patterns of SINEs, accumulating in gene-rich regions where other interspersed repeats are scarce, is apparently a conserved feature, independent of the primary sequence of the SINE… (Italics mine.)
Whatever the mystery cause is, it plucked out the species-specific SINEs from the junkety-junk LINE regions, and piled them high around the “twenty-five thousand genes” of the mouse and rat. Or it directed the SINEs to rain down on the gene-rich regions and in much lesser amounts elsewhere. This contradicts our selection hypothesis, unless the SINEs are doing something important in and around those protein-coding regions. But since so much ink has been spilled arguing that nothing of the sort is the case–these are junk elements, even harmful–we must turn to some other factor.
Reaching into the BioLogos box, we now pull out “genetic drift.” Neutral evolution means that a mutation–regardless of whether it is beneficial, neutral, or negative–can become fixed or lost in a lineage solely by chance. With respect to a SINE insertion, its persistence in a lineage would have to be a genetic coin toss: If heads, the SINE stays in a site; if tails, it is lost. So for a pure neutralist model to account for the graphs we have seen, ~300,000 random mutation events in the mouse have to match, somehow, the ~300,000 random mutation events in the rat.
What are the odds of that?
Thinking Outside the Box
Like the imaginary scientist trying to make sense of the far-from-random lunar evidence that Lunarlogos glossed over, I think we have to look elsewhere for the mystery cause of equivalent SINE patterns in the mouse and rat genomes. But where? A technical term was used in the sentence that I quoted above that you may have missed. I will highlight it for you:
“The cause of the unusual distribution patterns of SINEs, accumulating in gene-rich regions where other interspersed repeats are scarce, is apparently a conserved feature, independent of the primary sequence of the SINE and effective over regions smaller than isochores.”
Ever heard of “isochores”? Well, they are to DNA sequence organization along a chromosome what mountains and valleys are to a continent. Imagine buying a book about the geographical features of Africa and not finding a single word about Mount Kilimanjaro or the Great Rift Valley.
Imagine finding instead a lot of musings about what God couldn’t or wouldn’t have done with Africa. What would you think about such a book?
Well, turn to the index of Francis Collins’s The Language of God, or John Avise’s Inside the Human Genome: A Case for Non-Intelligent Design, and look for “isochore.” You won’t find it.
Isochores might provide a clue to cause of the mystery signal, but the cause–whatever it may be–is outside the BioLogos box.
Reference
1 Rat Genome Sequencing Project Consortium. 2004. Genome sequence of the Brown Norway rat yields insights into mammalian evolution. Nature 428: 493-521.