 Intelligent Design
Intelligent Design
The Machine that Fuels ATP Synthase
Why do you need oxygen to breathe? Oxygen actually plays a secondary role in the amazing process of respiration. What you really need are protons (hydrogen atoms). For every proton captured from your food, there’s an electron needing proper disposal. Oxygen is just an electron receptor at the end of a long chain of processes, driven by molecular machines, that captures protons for fuel. The machines translocate the protons across a membrane, creating a pool of protons that enter the ATP synthase rotor and make it turn (see our animation, “ATP Synthase: The Power Plant of the Cell“). In a sense, the whole job of respiration is to set up a proton gradient in the mitochondrial membrane to serve as fuel for ATP synthase.
A bit of background: The protons turn the rotor in ATP synthase like a carousel or waterwheel. This, in turn, rotates a camshaft to mechanically force ADP and phosphate into ATP in the catalytic center of the motor. The energy from your food (or from sunlight in plants) thus transforms chemical energy to electrical energy to mechanical energy and, finally, to another form of chemical energy. Most of the other processes in the cell use ATP for their energy.
We’ve heard of the mitochondrion as the “powerhouse” of the eukaryotic cell. That’s because it creates the ATP to power everything else. Along its folded inner membranes, called cristae, molecular machines pump protons to one side of the inner membrane where they can be channeled into the “turbines” of ATP synthase. In plants, chloroplasts serve this function, capturing energy from sunlight. Bacteria have the same basic machinery in their inner cell membranes. Since we are eukaryotes, let’s look at what’s going on in our mitochondria, where thousands of molecular machines are working 24/7 to set up the proton gradient, a literal “voltage” to run your motors.
The first of those machines has a cumbersome name, NADH:ubiquinone oxidoreductase (sometimes NADH dehydrogenase). We can use its nickname “Complex I” for convenience. It’s one of five “complexes” in the electron transport chain of respiration (also called oxidative phosphorylation), ending with ATP synthase as Complex V. The last of the molecular machines to be elucidated, Complex I has just been described in unprecedented detail by scientists from the molecular biology laboratory at Cambridge.
For many years, scientists knew the general function of Complex I. Its job is to generate four protons for the proton gradient from each input. It does this by taking electrons from NAD (nicotinamide adenine dinucleotide), a sugar phosphate first described in 1906. The reduced form is called NADH.
Now for some terminology: For historical reasons, removing electrons is called “oxidation”, and donating them is called “reduction.” Together, these are abbreviated “redox” reactions. But since negative electrons and positive protons are involved, it might help to think of ‘reduction’ as reducing the number of protons. Oxidizing a molecule leaves it with fewer electrons, resulting in a positive charge — i.e., with extra protons. Reducing a molecule leaves it with a negative charge, or a reduced number of protons. A proton is the same as a hydrogen ion (H+).
The docking site of Complex I oxidizes NADH to NAD+, passing two captured electrons to a cofactor called ubiquinone. In the process, by passing the electrons through a series of iron-sulfur clusters (Fe-S), the machine pumps four protons through the inner mitochondrial membrane, contributing about 40 percent of the proton gradient needed by ATP synthase. You can watch a simplified animation from NDSU Virtual Cell showing how the complexes move electrons and protons around.
The mitochondrion is where the well-known “citric acid cycle” takes place. Students often hear about the chemistry of life, but not as often about the mechanics. They learn how energy from food is transferred through various molecules to make ATP as we breathe in oxygen and breathe out carbon dioxide and water vapor. That’s great to know, but what is more fascinating is how these reactions require machines with moving parts. Let’s see what scientists have discovered about Complex I, in terms of its structure and dynamics.
Complex I is a huge enzyme, one of the largest in the cell. In mammals, it has 14 core subunits and 31 “supernumerary” (fancy word for “extra”) subunits, adding up to a whopping mass of 980 kilodaltons (kDa). (A Dalton is about the mass of a hydrogen atom; technically, 1/12 the mass of a carbon atom.) Such high mass implies over 7,000 properly-sequenced amino acids. That’s one huge machine, considering the average size of an enzyme is about 300-400 amino acids. The bacterial Complex I, lacking many of the supernumerary subunits, is still gigantic, weighing in at 550 kDa.
In appearance, Complex I resembles a boot, with the ankle inside the mitochondrion and the sole anchored in the crista. NAD enters the ankle. The protons exit the sole into the inner membrane. But what actually goes on in this structure? Research published in <href=”#ref9″>Nature in 2010 suggested the possibility that the bacterial enzyme moves with an action resembling a piston.
The overall architecture of this large molecular machine is now clear. F-ATPase has been compared to a turbine. In a similar vein, complex I seems to resemble a steam engine, where the energy of the electron transfer is used to move a piston, which then drives, instead of wheels, a set of discontinuous helices. The full mechanistic details remain to be clarified by atomic structures of the membrane domain and the entire complex.
A subsequent paper in <href=”#ref6″>Nature in 2014 called the piston-like motion into question, at least for the mammalian version, but it did not rule out smaller-scale motions (called “conformational changes” in the literature).
Now, using cryo-electron microscopy, the Cambridge team has described all 45 subunits of Complex I from bovine mitochondria. In Nature, they mention having found moving parts:
We have located and modelled all 45 subunits, including the 31 supernumerary subunits, to provide the entire structure of the mammalian complex. Computational sorting of the particles identified different structural classes, related by subtle domain movements, which reveal conformationally dynamic regions and match biochemical descriptions of the ‘active-to-de-active’ enzyme transition that occurs during hypoxia. Our structures therefore provide a foundation for understanding complex I assembly and the effects of mutations that cause clinically relevant complex I dysfunctions, give insights into the structural and functional roles of the supernumerary subunits and reveal new information on the mechanism and regulation of catalysis.
The supernumerary subunits “are central to the structure, stability and assembly of the complex, and some also have regulatory or independent metabolic roles,” they say. Some of them may serve a role in anchoring the machine to the membrane. That makes sense if the machine is vibrating from moving parts. What do the dynamic regions do? Later in the paper, they explain:
The two states of mammalian complex I described support the idea that dynamic, flexible regions at the hydrophilic-membrane domain interface are important for coupling ubiquinone reduction to proton translocation.
They go into detail about additional movements in a chain reaction, concluding:
Thus, a cascade of events originating from the ubiquinone-binding cleft may couple ubiquinone reduction and protonation to proton translocation. Although all such mechanisms for complex I are currently hypothetical, cryoEM now provides a powerful tool to study individual trapped conformations or separate mixed states computationally in order to determine how conformational changes are initiated, coordinated and propagated.
Currently, biochemists are limited to catching snapshots of the action. In the future, will they be able to watch Complex I move in real time? That’s something to look forward to!
Students are likely to be much more interested in cell biology if they learn about molecular machines with moving parts. Who wants to memorize the chemical reactions in the citric acid cycle when you can watch rotors, pistons and pumps? That’s what really goes on. We are privileged to live in a time when these realities are coming to light.
The authors point out two other observations of interest for intelligent design. One is that mutations in these machines cause disease and death; they cannot tolerate much change, meaning that the specificity in the amino acid sequence is vital to the function. That’s why they say that the core machinery is “conserved from bacteria to humans.”
The other observation is that the machines have to be assembled to work in the first place. It’s like Scott Minnich’s comment in Unlocking the Mystery of Life that the assembly instructions for the bacterial flagellum are even more complex than the machine itself. A machine needs a plan (encoded in DNA). It needs materials that must be delivered to the right place at the right time, in the right quantities. The parts have to be assembled in a coordinated sequence. Each step requires inspection, so that the cell doesn’t waste time building something that won’t work. That’s true of Complex I and the entire factory of machines in the electron transport chain that make life possible. We see similar requirements in the construction of a house or manufacturing plant. It’s Undeniable that we compare these processes and intuitively understand that intelligent causes must have been at work in the design of molecular machines.
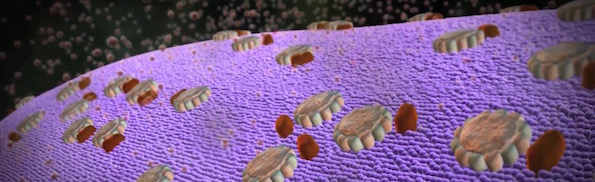
Let’s end with a look at one more level of organization. We mentioned cristae, the folds in mitochondrial membranes where these machines reside. A paper in the Proceedings of the National Academy of Sciences shows that the machines are arranged on the cristae in such a way as to maximize efficiency. In particular, the ATP synthase engines form V-shaped pairs, offset with respect to neighboring pairs so that their moving parts do not conflict but rather promote their respective operations. The spacing and angular displacement of the pairs, furthermore, results in the characteristic curvature of the cristae, which maximizes the proton gradient by creating local concentrations of protons aimed at the engines.
And this was found in Paramecium.
Image from: ATP Synthase: The Power Plant of the Cell.