 Evolution
Evolution
 Paleontology
Paleontology
Fossil Friday: The Mystery of the Frankenstein Dinosaur

This Fossil Friday we will look at a paleontological enigma that reveals a lot about the weaknesses of reconstructions of evolutionary trees and the phylogenetic classification of organisms, based on such shaky tree reconstructions. Paleobiologists use a methodology called cladistic analysis to reconstruct the assumed evolutionary relationships of fossil organisms. Of course, this cladistic methodology makes certain unproven assumptions such as the principle of parsimony and bifurcating (dichotomous) branching, even though nature is under no obligation to favor the most parsimonious trees or to avoid polytomous branchings or even network-like relationships (e.g., due to hybridization). But anyway, here is in simple words how cladistic tree reconstruction works in principle.
How It Is Done
The scientists compile a list of observable and relevant anatomical features of a fossil organism and possible fossil and living relatives. They construct a spread-sheet-like data matrix with different taxa as rows and different characters as columns. The fields of this data matrix allow for the coding of all character states for every taxon. These character states are usually coded as 0 for absent (or primitive / plesiomorphic state) and 1 for present (or derived / apomorphic state). Then a computer algorithm is used to find the most parsimonious branching pattern of a phylogenetic tree, which best explains the distribution of character states in the data matrix with the minimal number of necessary steps for evolutionary gains and losses. A so-called out-group taxon is included in the analysis, which is either a taxon that is presumed to be certainly more distantly related than all the other included taxa to each other, or is a hypothetical ancestor, which is coded with 0 for all character states. The tree is rooted in a way that this out-group represents the first branching of the tree. Only groups that include all branches of a node are recognized in the phylogenetic classification as monophyletic groups or clades, while grades of transitional series (paraphyletic groups) or polyphyletic groups that do not share a unique common ancestor are rejected as artificial groupings. This is why modern classifications no longer include taxa like invertebrates, or fish, or reptiles. The attribution of a fossil to a certain group must only be based on so-called shared derived characters (synapomorphies) but not on primitive similarities (symplesiomorphies) or convergent similarities (homoplasies).
So Far So Good
Now, given that we have a very extensive knowledge of most groups of dinosaurs, what would you expect if we find a new dinosaur that is almost completely and well preserved. Shouldn’t we be quite easily able to place it in the existing phylogenetic system based on the observed putative synapomorphies, thus the shared derived similarities with already known forms? Well, this works in happy Darwinian fantasy land of text books and the propaganda of popularizers of Darwinism like Richard Dawkins and Jerry Coyne, but the reality is up for a crude awakening. Often new discoveries show a confusing combination of characters that either overturn the previously preferred phylogenetic trees or require numerous ad hoc hypotheses to explain away the conflicting characters. Here is a very good example:
A few years ago, a new dinosaur genus and species, named Chilesaurus diegosuarezi, was described after a rather complete skeleton from the Late Jurassic of Chile (Novas et al. 2015). This dinosaur roamed Patagonia about 147-148 million years ago (Suárez et al. 2015). Actually, several isolated bones were already discovered and described previously, but erroneously considered to belong to different dinosaur taxa because they look so totally unrelated (Salgado et al. 2008, 2015). The original describers realized that the new dinosaur represents a quite enigmatic taxon with a bizarre combination of traits (Novas et al. 2015), and others called it “one of the most puzzling and intriguing dinosaurs ever discovered” (Barrett quoted in Rahim 2017). Initially, it was considered to be a derived theropod (Novas et al. 2015, Chimento et al. 2017, Lenin-Chávez et al. 2017, Cau 2018), thus a relative of the bipedal carnivorous dinosaurs, in spite of its spatulate dentition that clearly suggests a plant-eater (also see Lemonick 2015). It was even suggested that the forelimb structure foreshadows the acquisition of flight adaptations in avian theropods (Chimento et al. 2017). The very same year, a new study by Baron & Barrett (2017) suggested a very different affinity as basal ornithischian and a transitional ‘missing link’ between Ornithischia and Theropoda (University of Cambridge 2017). This was celebrated by media head lines around the globe as a final solution of the “mystery of the Frankenstein dinosaur” (Anonymous 2017, Geggel 2017, Gosh 2017, Rahim 2017).
One Year Later
However, just another year later a new study by Müller et al. (2018) disagreed and suggested that Chilesaurus was instead a basal representative of the sauropod lineage, but also cautioned that:
… these results demonstrate how search parameters, character scoring and taxon sampling could affect the phylogenetic position of C. diegosuarezi. Accordingly, our replication of Baron & Barrett’s [1] is compelling evidence that the phylogenetic status of C. diegosuarezi remains unstable and the mystery of this enigmatic dinosaur still remains unsolved.
Baron & Barrett (2018) defended their hypothesis, and subsequently, Müller & Dias-da-Silva (2019) changed their mind in a new study and now agreed that Chilesaurus is the basal-most member of Ornithischia, but again cautioned about unstable branches and “great uncertainty about the basic structure of the dinosaur family tree.”
A more recent study by Norman et al. (2022) recovered Chilesaurus as a highly derived ornithischian deeply nested in the family Heterodontosauridae. However, the authors also commented:
The ‘missing-link’ interpretation of its placement (Baron & Barrett, 2017) is incongruent chronologically (Late Jurassic) and evolutionarily, in the context of the acquisition of the fundamental ornithischian bauplan, but its curious opisthopubic pelvic anatomy may point toward either iterative (the repeated theropodan acquisition of opisthopuby) or atavistic anatomical phenomena. It is clear that the relationship of Chilesaurus in Dinosauria needs clarification. Most recently, Federico Agnolín (pers. comm., 25 April 2022) has reaffirmed the theropod affinities of Chilesaurus on the basis of the presence of pleurocoels, complex laminae on the cervical vertebrae, the shape of the ilium and carpal anatomy.
Baron (2022) admitted that “despite these recent fluctuations, the original placement within Theropoda remains the most prevalent phylogenetic hypothesis” and that the results of his new “analysis do not by any means end the debate”, but claimed that it “produced results that again suggest that Chilesaurus could be an early diverging member of Ornithischia.” I suggest our confidence in this “could” should not be too high.
A Complete Revolution?
Of course, it is also interesting in this context that the working group of Matthew Baron and Paul Barrett published a sensational new Ornithoscelida-hypothesis of dinosaur relationships (Baron et al. 2017), which suggested a complete revolution of the traditional division of dinosaurs into those with lizard-like hips (Saurischia) and those with bird-like hips (Ornithischia). Needless to say, the new phylogeny is still highly controversial and was quickly met with some support (Cau 2018, Müller & Dias-da-Silva 2019) but also with strong skepticism and criticism by proponents of the classical hypothesis (Müller & Garcia 2020, Novas et al. 2021), which ruled dinosaur classification since 1888. Curiously, the feathered non-avian theropods and birds do not belong to the clade with bird-like hips in either hypothesis, which is quite telling in itself. Actually, the disagreement is even worse, as was convincingly shown by Norman et al. (2022: fig. 1), who documented that basically every possible topology of the relationships between the three major groups of dinosaurs has be advocated by some modern experts (see the image below).
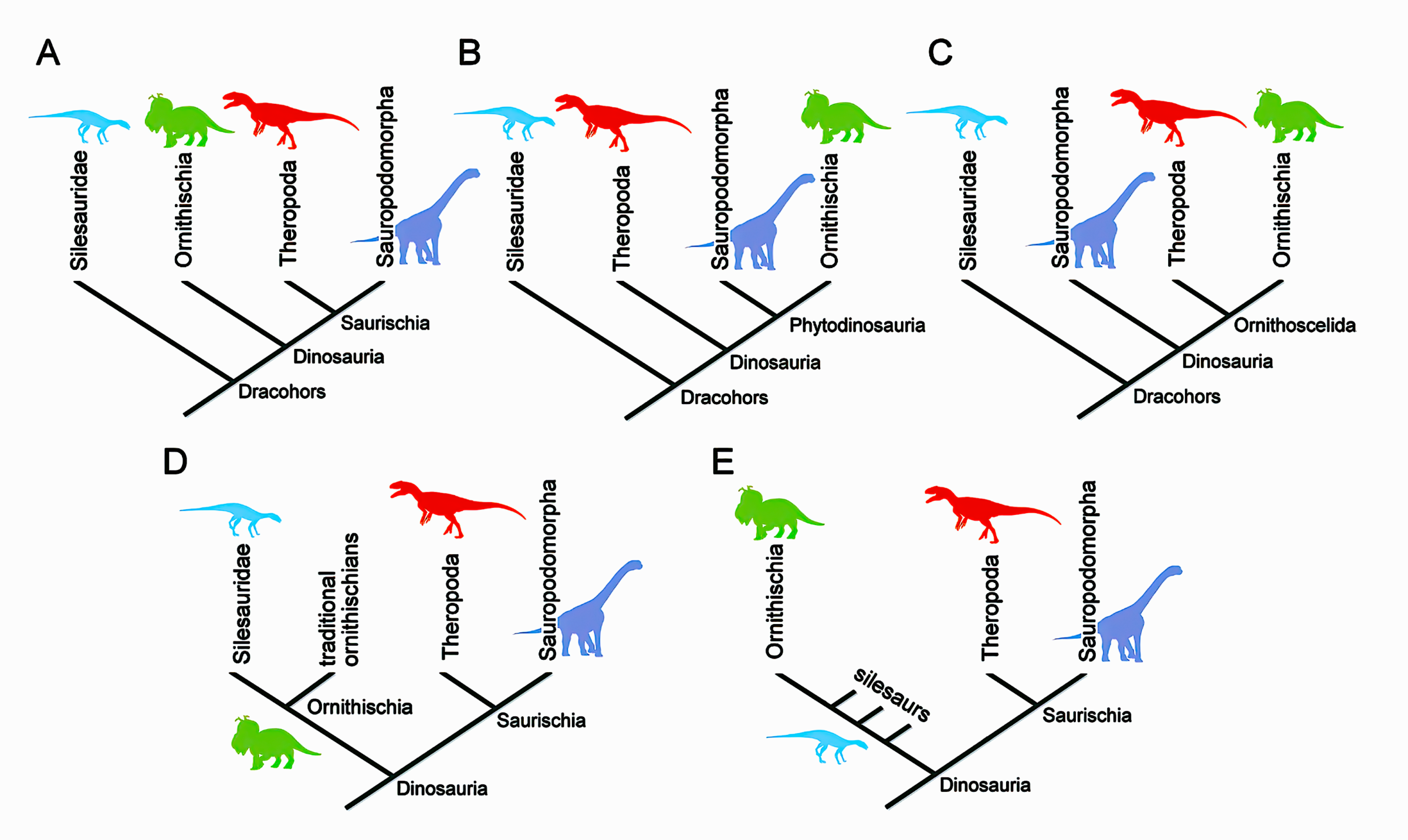
The Collapsing Tree Challenge
This mess is yet another confirmation of my collapsing tree challenge to neo-Darwinists (Bechly 2024). Phylogenetics very much looks like a pseudoscience (such as astrology or homeopathy) with a very sophisticated methodology, which fails to produce any consistent results. Different studies based on different data do not converge on one true tree of life. When Darwin sketched his famous first tree of life in his notebook, he titled it “I think.” Maybe he would think again today. We definitely should!
References
- Anonymous 2017. Monster mash: does the Frankenstein dinosaur solve the mystery of the Jurassic family tree? The Guardian August 16, 2017. https://www.theguardian.com/science/shortcuts/2017/aug/16/frankenstein-dinosaur-chilesaurus-diegosuarezi-mystery-jurassic-family-tree
- Baron MG 2022. The effect of character and outgroup choice on the phylogenetic position of the Jurassic dinosaur Chilesaurus diegosaurezi. Palaeoworld 33(1), 142–151. DOI: https://doi.org/10.1016/j.palwor.2022.12.001
- Baron MG & Barrett PM 2017. A dinosaur missing-link? Chilesaurus and the early evolution of ornithischian dinosaurs. Biology Letters 13(8):20170220, 1–5. DOI: https://doi.org/10.1098/rsbl.2017.0220
- Baron MG, Norman DB & Barrett PM 2017. A new hypothesis of dinosaur relationships and early dinosaur evolution. Nature 543(7646), 501–506. DOI: https://doi.org/10.1038/nature21700
- Baron MG & Barrett PM 2018. Support for the placement of Chilesaurus within Ornithischia: a reply to Müller et al. Biology Letters 14(3):20180002, 1–2. DOI: https://doi.org/10.1098/rsbl.2018.0002
- Bechly 6 2024. Fossil Friday: Three Modern Scientific Challenges to the Causal Adequacy of Darwinian Explanations. Evolution News May 17, 2024. https://evolutionnews.org/2024/05/fossil-friday-three-modern-scientific-challenges-to-the-causal-adequacy-of-darwinian-explanations/
- Cau A 2018. The assembly of the avian body plan: a 160-million-year long process. Bollettino della Società Paleontologica Italiana 57(1), 1–25. https://www.researchgate.net/publication/324941372 [DOI is broken]
- Chimento NR, Agnolin FL, Novas FE et al. 2017. Forelimb Posture in Chilesaurus diegosuarezi (Dinosauria, Theropoda) and Its Behavioral and Phylogenetic Implications. Ameghiniana 54(5), 567–575. DOI: https://doi.org/10.5710/amgh.11.06.2017.3088
- Geggel L 2017. This Enigmatic Dinosaur May Be the Missing Link in an Evolution Mystery. LiveScience August 16, 2017. https://www.livescience.com/60150-dinosaur-family-tree-missing-link.html
- Ghosh P 2017. ‘Frankenstein dinosaur’ mystery solved. BBC August 16, 2017. https://www.bbc.com/news/science-environment-40890714
- Lemonick MD 2015. T. rex’s Oddball Vegetarian Cousin Discovered. National Geographic April 27, 2015. https://web.archive.org/web/20150429185152/http://news.nationalgeographic.com/2015/04/150427-theropod-dinosaur-vegetarian-rex-science/
- Lenin-Chávez C, Ocampo-Cornejo P & Manzanero E 2017. Depicting Chilesaurus diegosuarezi (Dinosauria, Theropoda). Conference poster. https://www.researchgate.net/publication/376409197
- Müller RT & Dias-da-Silva S 2019. Taxon sample and character coding deeply impact unstable branches in phylogenetic trees of dinosaurs. Historical Biology 31(8), 1089–1092. DOI: https://doi.org/10.1080/08912963.2017.1418341
- Müller RT & Garcia MS 2020. A paraphyletic ‘Silesauridae’ as an alternative hypothesis for the initial radiation of ornithischian dinosaurs. Biology Letters 16(8):20200417, 1–5. DOI: https://doi.org/10.1098/rsbl.2020.0417
- Müller RT, Augusto Pretto F, Kerber L, Silva-Neves E & Dias-da-Silva S 2018. Comment on ‘A dinosaur missing-link? Chilesaurus and the early evolution of ornithischian dinosaurs’. Biology Letters 14(3):20170581, 1–2. DOI: https://doi.org/10.1098/rsbl.2017.0581
- Norman DB, Baron MG, Garcia MS & Müller RT 2022. Taxonomic, palaeobiological and evolutionary implications of a phylogenetic hypothesis for Ornithischia (Archosauria: Dinosauria). Zoological Journal of the Linnean Society 196(4), 1273–1309. DOI: https://doi.org/10.1093/zoolinnean/zlac062
- Novas F, Salgado L, Suárez M et al. 2015. An enigmatic plant-eating theropod from the Late Jurassic period of Chile. Nature 522(7556), 331–334. DOI: https://doi.org/10.1038/nature14307
- Novas FE, Agnolin FL, Ezcurra MD, Müller RT, Martinelli A & Langer M 2021. Review of the fossil record of early dinosaurs from South America, and its phylogenetic implications. Journal of South American Earth Sciences 10:103341. DOI: https://doi.org/10.1016/j.jsames.2021.103341
- Rahim Z 2017. ‘Frankenstein dinosaur’ enigma solved. CNN August 16, 2017. https://edition.cnn.com/2017/08/16/health/frankenstein-dinosaur-chilesaurus/index.html
- Salgado L, De La Cruz R, Suárez M, Gasparini Z & Fernández M 2008. First Late Jurassic dinosaur bones from Chile. Journal of Vertebrate Paleontology 28(2), 529–534. DOI: https://doi.org/10.1671/0272-4634(2008)28[529:FLJDBF]2.0.CO;2
- Salgado L, Novas FE, Suarez M, De La Cruz R, Isasi M, Rubilar-Rogers D & Vargas A 2015. Late Jurassic Sauropods in Chilean Patagonia. Ameghiniana 52(4), 418–429. DOI: https://doi.org/10.5710/amgh.07.05.2015.2883
- Suárez M, De La Cruz R, Fanning M, Novas F & Salgado L 2015. Tithonian age of dinosaur fossils in central Patagonian, Chile: U–Pb SHRIMP geochronology. International Journal of Earth Sciences 105(8), 2273–2284. DOI: https://doi.org/10.1007/s00531-015-1287-7
- University of Cambridge 2017. Study identifies dinosaur ‘missing link’. University of Cambridge press release August 16, 2017. https://www.cam.ac.uk/research/news/study-identifies-dinosaur-missing-link
