 Evolution
Evolution
Namacalathus, an Ediacaran Lophophorate Animal?
I have been writing a series of articles on alleged Ediacaran animals that have been postulated as precursors of the Cambrian explosion. Today and Friday we will have a look at Namacalathus, a problematic fossil organism from the Ediacaran Small Shelly Fauna (Hofmann & Mountjoy 2001, Bengtson 2004, Zhuralev et al. 2012). I will append my References section tomorrow.
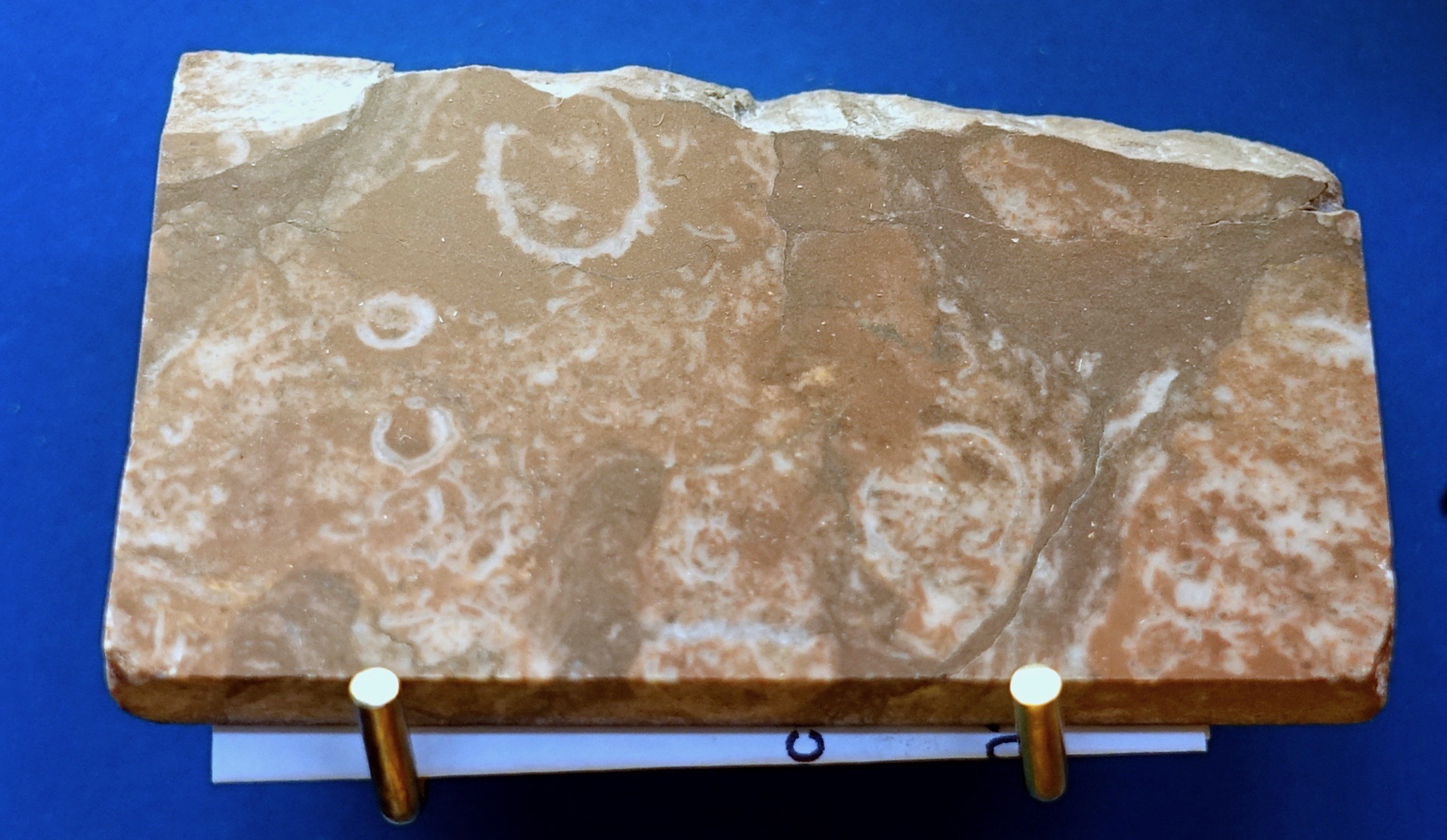
The Ediacaran Small Shelly Fauna is very different from the Cambrian Small Shelly Fauna (Grotzinger et al. 2000). Apart from Namacalathus it mainly includes Cloudina (the two are pictured above) and its relatives (cloudinomorphs) as well as the genus Namapoikia. These three Ediacaran calcareous taxa are generally considered to be the oldest known macroscopic biomineralized (skeletal/shelly) organisms. I have discussed Cloudina and Namapoikia in previous articles (Bechly 2020a, 2020b). These small shelly fossils are never found together with the typical Ediacaran organisms like Dickinsonia or the frond-like taxa. As Butler (2015) remarked: “Interestingly, Ediacaran organisms and these early shelly organisms are never found together, but they can be found in alternating layers, perhaps indicating that they preferred different environments.”
Overlooked for Years
For several years the weird Namacalathus fossils were overlooked and considered as deformed specimens of Cloudina (e.g., Grant 1990: fig. 8B; see Zhuravlev et al. 2012), with which they are generally found in association. Namacalathus hermanastes was first recognized and described by Grotzinger et al. (2000) from the microbial reefs of the 548-541 million years old Nama Group in southern Namibia (also see Grotzinger et al. 2005, Wood 2011, and Penny et al. 2017). Namacalathus has since also been found in the Kolodzha and Raiga Formations of West Siberia and the Anastas’ino Formation of the Altay-Sayan Foldbelt in Siberia (Kontorovich et al. 2008, 2009, Terleev et al. 2011, Zhuravlev et al. 2012, Grazhdankin et al. 2015), the Birba Formation of Oman (Amthor et al. 2003), the Byng Formation of British Columbia in Canada (Hofmann & Mountjoy 2001), and the Itapucumi Group of Paraguay (Warren et al. 2017). Some material from the Vendian of Siberia was considered as a new and much smaller species of the same genus by Kontorovich et al. (2008) but it was not formally named. The wide distribution of Namacalathus suggests dispersal with a planktonic larval stage (Penny et al. 2017).
A Goblet-Shaped Organism
Namacalathus abruptly appeared in the terminal Ediacaran 548.8 million years ago, without any putative ancestors in older strata, and then globally disappeared likewise abruptly at the Ediacaran-Cambrian boundary about 542 million years ago (Amthor et al. 2003, Zhuralev et al. 2012). Like Cloudina it is therefore considered as index fossil for the terminal Ediacaran era (Kontorovich et al. 2008).

{kind=link}
Namacalathus hermanastes is a 5-50 mm long and 2-35 mm wide, goblet-shaped organism with a hollow stalk carrying a hollow bulbous cup that has a wide apical opening at the top and a series of about six (5-7) openings around the side of the cup. Namacalathus was a solitary organism but is found in dense aggregations in fills between stromatolites and thrombolite reefs. The walls of its skeleton are very thin (ca. 100 μm thick), triple-layered with two thin inner and outer layers encasing a thicker inner layer comprising irregularly arranged rod-like crystals. The skeletons originally were aragonitic rather than calcitic (Wood 2011, Pruss et al. 2018), and later underwent diagenetic conversion into high-magnesium calcite, which is probably related to a different seawater chemistry in the Ediacaran and earliest Cambrian eras (Porter 2007, Zhuravlev & Wood 2008). Brasier et al. (2011) proposed that living Namacalathus could have been unmineralized and that its fossils are based on “tufa-like” abiogenic encrustation, but this was refuted by other studies (Zhuravlev et al. 2012).
Namacalathus were environmental generalists (Penny et al. 2017) and probably lived as sessile benthic filter-feeders or suspension-feeders on the seafloor, where they were attached to the microbial-algal carpets with a basal unmineralized holdfast at the lower end of the stalk. It is still controversial if aggregations of Namacalathus and Cloudina did (Wood & Curtis 2015, Wood 2011, 2017) or did not (Grotzinger et al. 2005) participate in substantial way to reef-building. The only lightly mineralized calcitic fossils cannot be separated from the calcitic stone matrix with conventional techniques, so that the three-dimensional shape had to be digitally reconstructed with computed tomographic analysis (Grotzinger et al. 2000, Watters & Grotzinger 2001). An interactive 3D-model of Namacalathus is available on the website of the Oxford University Museum of Natural History.
Putative Cnidarian
Namacalathus was originally described as a putative cnidarian by Grotzinger et al. (2000), who explicitly emphasized that these fossil organisms “do not appear to be closely related to the well-skeletonized bilaterian animals that radiated in younger oceans” and “Namacalathus hermanastes and other calcified Nama fossils are morphologically, and perhaps phylogenetically, distinct from Paleozoic skeleton formers.” Most subsequent authors (e.g., Wood 2011, Kouchinsky et al. 2012) also considered Namacalathus as calcareous cnidariomorphs.
Grotzinger et al. (2000) alternatively considered an affinity with sponges, especially the superficially similar Cambrian archeocyathans. Such a poriferan relationship was dismissed by Antcliffe et al. (2014) in their large revision of alleged Precambrian sponges. That is because apart from the overall hollow shape with an aboral opening there are no specific similarities with sponges, and the most distinctive sponge characters like pores and spicules are absent. In particular, the lateral ring of six openings is not only inconsistent with a poriferan but even with a cnidarian affinity. That is why Grotzinger et al. (2000) suggested that this character is just a diagenetic artifact. This was refuted by later studies, while Landing et al. (2018) sniped: “It also seems rather peculiar that in a fossil with so few characters, that its assignment would attempt to explain away one of these very few characters as diagenetic because it is not easily explicable in the context of a preferred phylogenetic hypothesis.”
Grotzinger et al. (2000) excluded a possible affinity with calcified green or red algae, because no known living forms possess a similar goblet-shaped morphology. This was controverted by Antcliffe et al. (2014), who suggested that Namacalathus is most likely a calcareous alga and mention an example for goblet-shape among the Dasycladales algae. Landing et al. (2018) concluded:
As is the case in Cloudina, there are thousands of Namacalathus specimens and none preserve septa or mesenteries. Similarly, there is no evidence for intergamet communication, axial structures, peripheral dissepiments, mural pores, tunnels, connective tubules or any of the other features that would allow confident placement in the Cnidaria. Indeed, the calcareous “goblet on a stalk” morphology is not widely distributed among the Cnidaria, yet it is common among a range of algal groups which in our opinion have been dismissed far too easily from consideration. At the very least, it is controversial whether Namacalathus represents a fossil eumetazoan and claims for its coloniality are not compelling.
Even though Grotzinger et al. (2000) had dismissed the possibility of a protist nature, Seilacher et al. (2003) suggested that Namacalathus could be a rhizopod protist due to its small size and an apparent lack of accretionary growth. Wood (2011) suggested that it belongs to the diploblast grade, i.e., stem- or crown-group cnidarians or basal stem-group bilaterians but did not support this with any evidence.
A Sensational Recent Study
After a lophotrochozoan relationship was already suspected by Zhuravlev et al. (2012), more recently, a sensational study was published by Zhuravlev et al. (2015) in the prestigious Proceedings of the Royal Society. The authors identified Namacalathus as a triploblastic bilaterian eumetazoan animal of the clade Lophophorata, which also includes brachiopods and bryozoans. The press release by the University of Edinburgh (2015), which was widely distributed through popular outlets like Science Daily and Phys.org, said: “The find is the first to suggest the earliest complex animals on Earth – which may be related to many of today’s animal species – lived millions of years earlier than was previously known.”
Unsurprisingly, the findings were celebrated by the blogosphere (Anonymous 2015, Bauwens 2015) and by international media as a “big piece of animal evolutionary history” (Joel 2015). They were even advertised to kids as the “earliest ever animal,” as theNational Geographic claimed: “Many of today’s animals could count the Namacalathus as their ancestors. … The discovery has changed scientists’ belief that most major animal groups evolved during the Cambrian Period.” Indeed, one of the co-authors of the study, Amelia Penny from the University of Edinburgh, wrote a blog post (Penny 2015) in which she maintained:
Our findings support the idea that complex animal groups evolved during the Ediacaran, long before the diversification of life habits and morphologies in the Cambrian. This suggests that the Cambrian Explosion was not the result of the rapid origin of animals. … There is increasing evidence that the evolutionary radiation of animals was not a geologically instantaneous event, and instead commenced tens of millions of years earlier.
This is a mouthful indeed. Tomorrow let’s take a look and see if the attribution of Namacalathus to lophophorate animals really is well justified and undisputed. Or is it just another piece of overhyped soft science? We should expect some excellent research and extraordinary evidence because paleontologist Marc LaFlamme from the University of Toronto, who was not involved in the study, commented in the Washington Post that: “I don’t remember seeing such detailed work done with any other Ediacaran fossil. This is the study that is going to set the bar for anyone else looking at Ediacaran fossils preserved like these specimens” (Joel 2015). I fear I have to disagree with that.