 Evolution
Evolution
 Intelligent Design
Intelligent Design
 Paleontology
Paleontology
Fossil Friday: Purgatorius and the Abrupt Origin of Primates
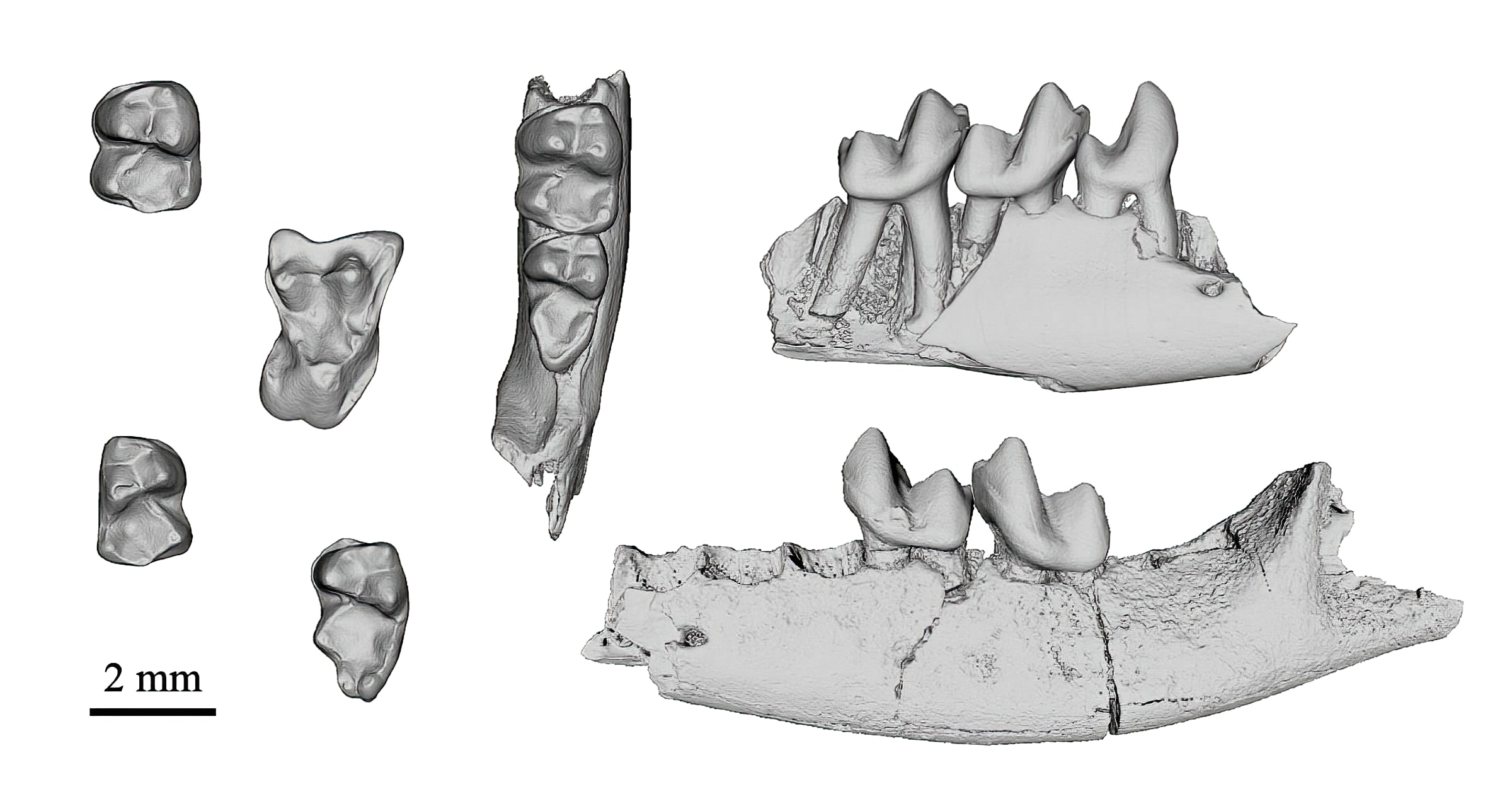
With this Fossil Friday I introduce a new series of articles on the age of origin of the various modern placental mammal orders. These orders are the higher categories of mammal systematics, which for example include groups like bats, rodents, primates, carnivores, sirenians, elephants, even-toed and odd-toed ungulates etc. This issue is quite relevant and important for ID research as it establishes the abrupt origin of all these groups after the end-Cretaceous mass extinction caused by the global consequences of the Chicxulub impact. This week we open this series with the origin of our very own order, Primates.
The bestselling author Stephen Baxter (2003) mused in the first chapter of his novel Evolution what it was like to be “Purga,” one of our remote primate ancestors, on the last day of the reign of the dinosaurs before the asteroid impact. He might have borrowed this idea from the popular Disney animation movie Dinosaur (2000), which featured lemur-like primates frolicking with dinosaurs until the cosmic cataclysm ended the edenic scenery. Is this just Hollywood fantasy like The Flintstones or is there some scientific support for such a picture? Indeed, a recent study was celebrated in media reports as suggesting that primates walked with dinosaurs (McKeever 2021). Let’s have a look.
The fossils featured today are dental remains of the small Paleogene mammal Purgatorius, which is generally considered to belong to the extinct order Plesiadapiformes as their oldest and most primitive representative (Van Valen 1994, Clemens 2004, Rose 2006, Fox & Scott 2011). Plesiadapiformes is a very diverse group of more than 140 named species in 50 genera and 11-12 families that lived between the Early Paleocene and the Late Eocene (Fleagle 2013, Silcox 2014, Silcox et al. 2017). The genus Purgatorius was named after the Purgatory Hill locality in Montana, where it was first described by Van Valen & Sloan (1965), but has also been found in contemporaneous outcrops in Canada (Fox & Scott 2011). Most of the finds are of Puercan age, which is a brief period of only 1 million years in the Early Paleocene (Lofgren et al. 2004). These tiny animals presumably resembled squirrels and certainly had an arboreal way of life (Fiegl 2012, Chester et al. 2015, 2017, 2019). They are not yet known by skeleton finds, but only by isolated teeth or jaw fragments and some ankle bones, which is a very common situation in fossil mammals and paleoanthropology.
A Single Tooth
Anyway, until recently the oldest fossil record of Purgatorius was from the Early Paleocene of North America. A single tooth from the Hell Creek Formation of Harbicht Hill in Montana was initially considered to be of Late Cretaceous origin, because it was found in the same deposit as Triceratops dinosaur remains (Van Valen & Sloan 1965), but it was later shown to be a Tertiary intrusion in sediments of mixed origin (Clemens 2004). Other alleged Cretaceous material from the Ravenscrag Formation and the Bug Creek Group was later re-dated to a Paleocene age (Lillegraven et al. 1979). Unfortunately, the long-refuted attribution to the Late Cetaceous is still considered in some more recent textbooks such as Fleagle (2013: 215).
Clemens (2004) suggested that the abundance of Purgatorius in the Puercan 2-3 of Montana is explained by a dispersal in the Early Paleocene 64.75-64.11 million years ago. Chester et al. (2015) described tarsal bones attributed to Purgatorius from late Puercan (65 mya) of Montana. A new study by Wilson Mantilla et al. (2021) described three new species of Purgatorius from this locality, which has been more precisely dated to be of Earliest Paleocene age (65.921 mya), only about 105-139 thousand years after the K/Pg boundary (McKeever 2021, Sanders 2021). They concluded that these oldest plesiadapiform fossils suggest “purgatoriids and, by extension, Pan-Primates, Euarchonta and Placentalia probably originated by the Late Cretaceous”. That would be a reasonable conclusion if and only if Purgatorius was a placental mammal and a stem primate. So, was the Disney movie history after all? Sorry, to disappoint any fans, but here comes the fly in the ointment.
Early Primates?
Initially, plesiadapiforms and Purgatorius were indeed considered as early primates (Van Valen & Sloan 1965, Clemens 1974, Szalay & Delson 1979, Van Valen 1994), mainly because they share certain arboreal adaptations (e.g., long fingers) and some similar dental features, even though they differed by still having claws and smaller lateral orbits. Furthermore, the dental similarities are only developed in some plesiadapiforms (Rose et al. 1994), so that Rose (1994) already found that “the detailed dental similarity must be convergent” and generally concluded:
“Although considerable evidence has been adduced to ally plesiadapiforms with primates, adapids with strepsirhines and anthropoids, and omomyids with tarsiers and haplorhines,much of it is based on either superficial resemblance, symplesiomorphy, or obvious convergence. Controversy persists because compelling evidence of these relationships, in the form of clear and significant synapomorphies, is still wanting. Opinion is increasing that Plesiadapiformes are not primates …”
Of course, a potential solution for this incongruent distribution of similarities could be a nesting of primates within a paraphyletic plesiadapiform grade, so that some plesiadapiform genera could be more closely related to primates than others. This was indeed suggested by the study of Bloch et al. (2007). However, as emphasized by Fleagle (2013):
“The scenario in which crown primates are nested within plesiadapiforms as the sister taxon of plesiadapoids involves an evolutionary reversal of increasing tooth number and dramatic reduction of the procumbent incisors that seems unlikely” (also see Godinot 2017).
Concerning the arboreal adaptations, more recent studies of living tree shrews have suggested that these adaptations already belonged to the archontan ground plan (Archonta are the supposed clade including tree shews, colugos, and primates) and thus do not suggest a uniquely primate relationship (Godinot 2017).
Considerable Scientific Debate
The significance of the similarities between plesiadapiforms and primates is still matter of considerable scientific debate to this day (e.g., Silcox et al. 2007 vs Soligo & Martin 2007). Already in the 1960-80s the consensus more and more shifted towards an exclusion of Plesiadapiformes from the order Primates (Hartwig 2002, Fleagle 2013). Rose (2006) is one of the few exceptions and tentatively accepted plesiadapiforms as early stem primates, but he readily admitted that:
“Plesiadapiforms, as well as euprimates, are usually traced back to early Paleocene Purgatorius, although no transitional forms leading to euprimates have been identified, and the source of Purgatorius itself is completely unknown …”
Most other authors instead considered Plesiadapiformes to be a separate order, which is either the sister group of primates, or of colugos (Dermoptera), or of both together (Kemp 2005, Silcox 2014, Godinot 2017). Kay et al. (1992) proposed a close relationship of plesiadapiforms and colugos based on the shared reduction of the internal carotid artery. Beard (1990, 1993) and McKenna & Bell (1997) came to the same result based on skeletal characters, and a recent cladistic analysis confirmed this position (Morse et al. 2019). Kemp (2005) commented in his standard textbook on The Origin & Evolution of Mammals that “Purgatorius is the earliest member of a diverse group, Plesiadapiformes, which many authors no longer believe to be primates”. But then a cladistic study by Bloch et al. (2007, 2016; also see Silcox 2001 and Bloch & Silcox 2006) indeed placed plesiadapiforms with primates and found no evidence supporting a dermopteran relationship. They suggested a divergence of the two groups about 62 million years ago, which they admitted to imply an euprimate ghost lineage of 7 million years. Chester et al. (2015, 2017, 2019) also recovered Purgatorius and other plesiadapiforms as early stem group representatives of primates. On the other hand, Ni et al. (2013, 2016) recovered plesiadapiforms as sister group to Dermoptera+Primates in their parsimony analyses and concluded “Plesiadapiforms, traditionally regarded as archaic primates, are not even stem primates, corroborating the now common practice of excluding plesiadapiforms from the order Primates”. Therefore, Godinot (2017) agreed with Hartwig (2002) that “there is no clear evidence that Plesiadapiformes are the closest sister group of the Euprimates”. In the most recent phylogenetic study, Plesiadapiformes and Purgatoriidae have been corroborated to be successively more closely related to a clade of colugos and primates (Seiffert et al. 2020), and excluded from the primate lineage with a 100% posterior probability in the Bayesian analysis.
Thus, neither plesiadapiforms in general nor Purgatorius in particular can be considered to be well-established fossil stem primates. The weak indirect evidence for a contemporaneous occurrence of early primates and dinosaurs evaporates into a mist of evolutionist speculation and storytelling. That said: in theory the evidence indeed could be consistent with the possibility that primate-like ancestors of the clade of colugos and primates may have lived in the latest stages of the age of dinosaurs close to the extinction event. Of course, there exists not a single fossil to empirically prove this speculation.
But It Gets Worse
There is substantial anatomical evidence, which suggests that Purgatoriusnot only has to be excluded from primates, but may not even be a crown group placental mammal. It rather seems to be related to enigmatic stem mammals like the “condylarthran” genus Protungulatum (Wible et al. 2007, 2009, Goswami et al. 2011, Halliday et al. 2015; also see Davies et al. 2017). Chester et al. (2015) mentioned this fact, but considered it as an artifact of taxon sampling, because their data matrix supported a primate affinity within placental mammals. This was mainly based on the ankle characters, so that it is surprising that the authors do not even consider the possibility that similarities between Purgatorius and primates could rather be based on convergent adaptation to an arboreal life. Apparently, Chester et al. were unaware of the concurrent study by Halliday et al. (2015), which they do not cite, while the Halliday paper included the evidence from Chester’s. The studies by Chester et al. (2017, 2019) and by Wilson Mantilla et al. (2021) can hardly be given such a benefit of doubt when they simply ignored this conflicting evidence and the crucial study by Halliday et al., which arguably represents the most important work on the affinities of enigmatic Paleocene mammals. Finding Purgatorius outside of placental mammals would have rendered Wilson Mantilla’s sensational story dead in the water. Honi soit qui mal y pense.
Since neither Purgatorius nor Plesiadapiformes seem to be stem primates, all the other plesiadapiform taxa, which have been suggested by some authors to rank among the earliest primates, also have to rejected as such. These include for example the following Paleocene genera and species (age ranges are based on the PaleoDB database at fossilworks.org):
- Carpolestes twelvemilensis (61.7-56.8 mya)
- Draconotus apertus (63.3-61.7 mya)
- Dryomomys millenius (61.7-56.8 mya)
- Elphidotarsius florencae (63.3-61.7 mya)
- Micromomys silvercouleei (61.7-50.3 mya)
- Nannodectes spp. (61.7-56.8 mya)
- Pronothodectes jepi and P. matthewi (63.3-61.7 mya)
- Russellodon haininense (66.043-61.7 mya) (De Bast & Smith 2017)
- Saxonella crepaturae (61.7-58.7 mya)
- Torrejonia wilsoni 62.4 mya (Chester et al. 2019)
But even if these plesiadapiform taxa would represent early stem primates, what they likely do not (see above), they would only document the presence of primates in the Early Paleocene but not prior to the dinosaur mass extinction at the K/Pg-boundary.
If Purgatorius and plesiadapiforms are not the earliest fossil primates, then which are the better supported candidates for this illustrious position?
A few teeth and a mandible fragment from the Late Paleocene of Morocco (about 58.7-55.8 mya) were described by Sigé et al. (1990) as Altiatlasius koulchii. The describers and many subsequent authors considered it to be the earliest euprimate (e.g., Rose 1994, Tabuce et al. 2004, Bloch et al. 2007, Godinot 2017), and maybe an omomyoid stem tarsier (Fleagle 2013) or even a simian (Godinot 1994, Beard 2006, Bajpai et al. 2008). However, the precise systematic position of Altiatlasius is very much disputed (Hartwig 2002, Fleagle 2013). Even a plesiadapiform relationship has been suggested (Hooker et al. 1999, Hartwig 2002) and has recently been supported by a cladistic analysis (Morse et al. 2019), which recovered Altiatlasius in a non-primate clade of plesiadapiforms and colugos. As in so many cases, phylogenetics proves to be a kind of guessing game rather than a hard science.
Another very old group of primitive primates is the extinct order Adapiformes (not to be confused with Plesiadapiformes), which is believed to be closer related to lemurs than to simians. Some of the oldest representatives are the notharctid genera Notharctus (50.3-40.4 mya), Cantius (50.3-40.4 mya), and Donrussellia (55.8-48.6 mya) from the Early Eocene of North America and France (Gingerich 1986, Hartwig 2002), as well as Marcgodinotius from the early Eocene (52 mya) of India (Bajpai et al. 2005). These early adapiforms arguably establish an appearance of the lemur lineage 55 million years ago.
Archicebus achilles was described by Ni et al. (2013) from the Earliest Eocene (55.8-54.8 mya) of Hubei Province in central China. It is a relatively complete skeleton of a tiny, tree-dwelling mammal, which was even smaller than the smallest living primate but had a monkey-like habitus and anatomy (Chen 2013, ESRF 2013). It is 7 million years older than all previously found early primate skeletons and was thoroughly studied with X-ray synchrotron tomography. It proved to be a very basal tarsiiform and thus closer related to simians (“monkeys”) than to lemurs (Fleagle 2013).
The Eocene genus Teilhardina, which is known in several different species from North America and Eurasia, belongs to the extinct family Omomyidae (Bown 1976, Rose 2006). This family is generally attributed to the stem group of tarsiers (Kemp 2005), or more rarely to the common stem of tarsiers and simians (Fleagle 2013, Morse et al. 2019) suggesting an Asian origin of this clade (Ni et al. 2005). These animals may have looked similar to modern bush babies. Teilhardina asiatica was discovered in the Hunan Province of China and dated to an earliest Eocene age of 54.97 mya (Ni et al. 2004). The fossil is a partial skull with complete dentition, which is a quite rare condition for early primate fossils. The closely related species Teilhardina magnoliana was found in the earliest Eocene Tuscahoma Formation from the Gulf Coastal Plain in North America (Beard 2008). Beard suggested that its ancestor must have crossed the land bridge connecting Siberia and Alaska more than 55.8 million years ago (Nickerson 2008), thus somewhat earlier than believed before (Smithet al. 2006). New material of Teilhardina brandti, which was originally described by Gingerich (1993) as oldest omomyid from North America, also is about 55.8 million years old (Rose et al. 2011, Boyer et al. 2018, Hoose 2018).
Teilhardina and Archicebus are the oldest well-dated primates, while all others suffer from a relatively wide range of uncertainty in their dating that could make them significantly younger. Therefore, these two genera are best supported by the evidence as earliest fossil record of primates and also establish the presence of the tarsier lineage about 55 million years ago.
Other Contenders
Some of those other contenders, especially for the position as oldest simians (Anthropoidea), are the following:
The genera Azibius and Algeripithecus are small-bodied primates from the Eocene of Algeria. Azibius was originally described by Sudre (1975) and considered to be a plesiadapiform (“paromomyid”), then recognized as adapid primate by Gingerich (1976), again transferred to Plesiadapiformes by Tabuce et al. (2004) which was questioned by Rose (2006), and yet again reinstated as a lemuriform primate (Tabuce et al. 2009). Algeripithecuswas originally described by Godinot & Mahboubi (1992) as earliest known simian, which was widely accepted and considered as support for an African origin of simians (Godinot 1994). A more recent evaluation of the age of the Glib Zegdou Formation in Algeria suggests an early-middle Eocene age of 49-45 mya (Coster et al. 2012), which would support this ranking. Rose (2006) commented on Algeripithecus that its “significance will remain moot until more complete evidence is found.” A few years later, Marivaux et al. (2011) studied the talus bone and concluded that Algeripithecus is not a simian at all but was closely related to Azibius and belonged to the strepsirrhine branch of primates that also includes lemurs. This was strongly confirmed by the more recent study of Tabuce et al. (2009), who also emphasized that this strongly challenges the role of Africa as the ancestral homeland for simians.
This fate reminds of the case of Darwinius marsillae from the Middle Eocene oil shale (ca. 47 mya according to Franzen 2005) of the Messel pit in Germany. See my previous Fossil Friday article (Bechly 2022) about the remarkable story of this fossil, which was nick-named “Ida” and heavily overhyped as one of the oldest simian fossils and an important “missing link”, only to be quickly revealed to be just another early relative of lemurs.
The currently oldest fossil record of simians is the extinct family Eosimiidae. It is based on the fragmentary remains of Eosimias sinensis described by Beard et al. (1994) from the Middle Eocene of Jiangsu Province in China. Its age was estimated to be about 45 mya, but unfortunately no radiometric dates were available for this deposit. The simian and even primate affinity of Eosimias was disputed by several other experts (e.g., Godinot 1994), but the discovery of better material of a second species of Eosimias confirmed its simian relationship (Beard et al. 1996, Beard & Wang 2004, Rose 2006), even though some experts still remain skeptical (e.g., Godinot 2017). The Middle Eocene age would be about 10 million years younger than the oldest fossil record for stem tarsiers. Evolutionists had to explain away this inconsistency with a so-called ghost lineage of undocumented existence, because simians of course have to be as old as their sister group tarsiers. However, unlike many other such cases this particular problem got solved by the new discovery of another genus of Eosimiidae, which was described as Anthrasimias by Bajpai et al. (2008) from the Early Eocene (55-54 mya) of India. Thus the first appearance of the simian lineage can also be dated to about 55 million years ago.
An Abrupt Appearance
We can conclude from all the mentioned up-to-date research that the placental mammal order of Primates appears abruptly in the fossil record of the Eocene about 55-56 million years ago, during the hothouse climate of the Paleocene-Eocene Thermal Maximum (PETM). This just confirms what Rose (1994) already had concluded in his seminal review article on the earliest primates:
“Undisputed primates appear suddenly in the Holarctic fossil record at the beginning of the Eocene, approximately 55 million years ago.”
The fact that almost thirty years of great progress in primate paleontology did not change this result provides some confidence that it is not an artifact of an incomplete fossil record, but is here to stay. Primates not only appeared suddenly, but their different subgroups of lemurs, tarsier, and simians all appeared at about the same time. Primates never shared the planet with dinosaurs, even if Hollywood and National Geographic want to sell you a different story. We will see in subsequent articles in this series that such an abrupt appearance in a narrow window of time of the Paleogene period represents a consistent pattern found in all the placental mammal orders. Such a saltational pattern contradicts Darwinian gradualist expectations and is better explained by pulses of new information infused into the system. Even Darwinists implicitly admit this when they say that “eutherians experienced elevated evolutionary rates in the immediate aftermath of the Cretaceous–Palaeogene mass extinction” (Halliday et al. 2016). Unlike ID theorists, they have no plausible explanation of how a meteorite impact produced the genetic information for an explosive diversification of placental mammals.
References
- Bajpai S, Kapur VV, Thewissen JGM, Das DP, Tiwari BN, Sharma R & Saravanan N 2005. Early Eocene primates from Vastan Lignite mine, Gujarat, western India. Journal of the Palaeontological Society of India 50(2), 43–45. http://palaeontologicalsociety.in/vol50_2.php
- Bajpai S, Kay RF, Williams BA, Das DP, Kapur VV & Tiwari BN 2008. The oldest Asian record of Anthropoidea. PNAS 105(32), 11093–11098. DOI: https://doi.org/10.1073/pnas.0804159105
- Baxter S 2003. Evolution. Del Rey, New York (NY), 592 pp.
- Beard KC 1990. Gliding behaviour and palaeoecology of the alleged primate family Paromomyidae (Mammalia, Dermoptera). Nature 345(6273), 340–341. DOI: https://doi.org/10.1038/345340a0
- Beard KC 1993. Phylogenetic systematics of the Primatomorpha, with special reference to Dermoptera. pp. 129–150 in: Szalay FS, Novacek MJ & McKenna MC (eds.). Mammal Phylogeny: Placentals. Springer-Verlag, Berlin (DE), xi+321 pp.
- Beard C 2004. The Hunt for the Dawn Monkey. University of California Press, Berkeley (CA), 348 pp.
- Beard KC 2006. Mammalian Biogeography and Anthropoid Origins. pp. 439–467 in: Lehman SM & Fleagle JG (eds). Primate Biogeography: Progress and Prospects. Springer, New York (NY), xii+536 pp. DOI: https://doi.org/10.1007/0-387-31710-4_15
- Beard KC 2008. The oldest North American primate and mammalian biogeography during the Paleocene–Eocene Thermal Maximum. PNAS 105(10), 3815–3818. DOI: https://doi.org/10.1073/pnas.0710180105
- Beard KC & Wang J 2004. The eosimiid primates (Anthropoidea) of the Heti Formation, Yuanqu Basin, Shanxi and Henan Provinces, People’s Republic of China. Journal of Human Evolution 46(4), 401–432. DOI: https://doi.org/10.1016/j.jhevol.2004.01.002
- Beard KC, Qi T, Dawson MR, Wang B & Li C 1994. A diverse new primate fauna from middle Eocene fissure-fillings in southeastern China. Nature 368 (6472), 604–609. DOI: https://doi.org/10.1038/368604a0
- Beard KC, Tong Y, Dawson MR, Wang J & Huang X 1996. Earliest Complete Dentition of an Anthropoid Primate from the Late Middle Eocene of Shanxi Province, China. Science272(5258), 82–85. DOI: https://doi.org/10.1126/science.272.5258.82
- Bechly G 2022. Fossil Friday: Darwinius, or How Wishful Thinking Makes a Missing Link. Evolution News September 23, 2022. https://evolutionnews.org/2022/09/fossil-friday-darwinius-or-how-wishful-thinking-makes-a-missing-link/
- Bloch JI & Silcox MT 2006. Cranial anatomy of Paleocene plesiadapiform Carpolestes simpsoni (Mammalia, Primates) using ultra high-resolution X-ray computed tomography, and the relationships of plesiadapiforms to Euprimates. Journal of Human Evolution 50(1), 1–35. DOI: https://doi.org/10.1016/j.jhevol.2005.06.009
- Bloch JI, Silcox MT, Boyer DM & Sargis EJ 2007. New Paleocene skeletons and the relationship of plesiadapiforms to crown-clade primates. PNAS 104(4), 1159–1164. DOI: https://doi.org/10.1073/pnas.0610579104
- Bloch JI, Chester SGB & Silcox MT 2016. Cranial anatomy of Paleogene Micromomyidae and implications for early primate evolution. Journal of Human Evolution 96, 58–81. DOI: https://doi.org/10.1016/j.jhevol.2016.04.001
- Bown TM 1976. Affinities of Teilhardina (Primates, Omomyidae), with description of a new species from North America. Folia Primatologica 25(1), 62–72. DOI: https://doi.org/10.1159/000155707
- Boyer DM, Maiolino SA, Holroyd PA, Morse PE & Bloch JI 2018. Oldest evidence for grooming claws in euprimates. Journal of Human Evolution 122, 1–22. DOI: https://doi.org/10.1016/j.jhevol.2018.03.010
- Chen N 2013. Oldest Known Fossil Primate Skeleton Found From the Eocene of Hubei, China. Chinese Academy of Sciences News June 6, 2013. https://english.cas.cn/newsroom/archive/news_archive/nu2013/201502/t20150216_140470.shtml
- Chester SGB, Bloch JI, Boyer DM & Clemens WA 2015. Oldest known euarchontan tarsals and affinities of Paleocene Purgatorius to Primates. PNAS 112(5), 1487–1492. DOI: https://doi.org/10.1073/pnas.1421707112
- Chester SGB, Williamson TE, Bloch JI, Silcox MT, Sargis EJ. 2017 Oldest skeleton of a plesiadapiform provides additional evidence for an exclusively arboreal radiation of stem primates in the Palaeocene. Royal Society Open Science 4:170329, 1–9. DOI: https://doi.org/10.1098/rsos.170329
- Chester SGB, Williamson TE, Silcox MT, Bloch JI & Sargis EJ 2019. Skeletal morphology of the early Paleocene plesiadapiform Torrejonia wilsoni (Euarchonta, Palaechthonidae). Journal of Human Evolution 128, 76–92. DOI: https://doi.org/10.1016/j.jhevol.2018.12.004
- Clemens WA 1974. Purgatorius, an Early Paromomyid Primate (Mammalia). Science184(4139), 903–905. DOI: https://doi.org/10.1126/science.184.4139.903
- Clemens WA 2004. Purgatorius (Plesiadapiformes, Primates?, Mammalia), a Paleocene immigrant into Northeastern Montana: stratigraphic occurrences and incisor proportions. In: Dawson MR & Lillegraven JA (eds). Fanfare for an Uncommon Paleontologist: Papers in Honor of Malcolm C. McKenna. Bulletin of the Carnegie Museum of Natural History 36, 3–13. DOI: https://doi.org/10.2992/0145-9058(2004)36[3:PPPMAP]2.0.CO;2
- Coster P, Benammi M, Mahboubi M, Tabuce R, Adaci M, Marivaux L, Bensalah M, Mahboubi S, Mahboubi A, Mebrouk F, Maameri C & Jaeger J-J 2012. Chronology of the Eocene continental deposits of Africa: Magnetostratigraphy and biostratigraphy of the El Kohol and Glib Zegdou Formations, Algeria. Geological Society of America Bulletin 124(9-10), 1590–1606. DOI: https://doi.org/10.1130/b30565.1
- Davies TW, Bell MA, Goswami A & Halliday TJD 2017. Completeness of the eutherian mammal fossil record and implications for reconstructing mammal evolution through the Cretaceous/Paleogene mass extinction. Paleobiology 43(4), 521–536. DOI: https://doi.org/10.1017/pab.2017.20
- De Bast E & Smith T 2017. The oldest Cenozoic mammal fauna of Europe: implication of the Hainin reference fauna for mammalian evolution and dispersals during the Paleocene. Journal of Systematic Palaeontology 15(9), 741–785. DOI: https://doi.org/10.1080/14772019.2016.1237582
- ESRF 2013. Oldest Known Primate skeleton casting a New light on the Origin of Anthropoids. ESRF Highlights 2013. https://www.esrf.fr/home/UsersAndScience/Publications/Highlights/highlights-2013/x-ray-imaging/im2.html
- Fiegl A 2012 . World’s Oldest Primate Was a Rodentlike Climber. National Geographic News October 26, 2012. https://www.nationalgeographic.com/science/article/121024-purgatorius-earliest-primate-evolution-science-squirrel
- Fleagle JG 2013. Primate Adaptation and Evolution. 3rd Edition. Academic Press, London (UK), 456 pp.
- Fox RC & Scott CS 2011. A New, Early Puercan (Earliest Paleocene) Species of Purgatorius (Plesiadapiformes, Primates) from Saskatchewan, Canada. Journal of Paleontology 85(3), 537–548. JSTOR: https://www.jstor.org/stable/23020189
- Franzen JL 2005. The implications of the numerical dating of the Messel fossil deposit (Eocene, Germany) for mammalian biochronology. Annales de Paléontologie 91(4), 329–335. DOI: https://doi.org/10.1016/j.annpal.2005.04.002
- Gingerich PD 1976. Cranial anatomy and evolution of early Tertiary Plesiadapidae (Mammalia, Primates). Papers on Paleontology 15, 1–141. https://hdl.handle.net/2027.42/48615
- Gingerich PD 1986. Early Eocene Cantius torresi – oldest primate of modern aspect from North America. Nature 320, 319–321. DOI: https://doi.org/10.1038/319319a0
- Gingerich PD 1993. Early Eocene Teilhardina brandti: Oldest Omomyid Primate from North America. Contributions of the Museum of Paleontology University of Michigan 28(13), 321–326. https://hdl.handle.net/2027.42/48723
- Godinot M 1994. Early North American primates and their significance for the origins of Simiformes (= Anthropoidea). pp. 235–295 in: Fleagle JG & Kay RF (eds). Anthropoid Origins. Plenum Press, New York (NY), 708 pp. DOI: https://doi.org/10.1007/978-1-4757-9197-6_10
- Godinot M 2017. Paleocene and Eocene Primates. pp. 1–9 in: Fuentes A (ed.). The International Encyclopedia of Primatology. John Wiley & Sons, Hoboken (NJ), 1608 pp. DOI: https://doi.org/10.1002/9781119179313.wbprim0331
- Godinot M & Mahboubi M 1992. Earliest known simian primate found in Algeria. Nature357(6376), 324–326. DOI: https://doi.org/10.1038/357324a0
- Goswami A, Prasad GVR, Upchurch P & Flynn JJ 2011. A radiation of arboreal basal eutherian mammals beginning in the Late Cretaceous of India. PNAS 108(39), 16333–16338. DOI: https://doi.org/10.1073/pnas.1108723108
- Halliday TJD, Upchurch P & Goswami A 2015. Resolving the relationships of Paleocene placental mammals. Biological Reviews 92(1), 521–550. DOI: https://doi.org/10.1111/brv.12242
- Halliday TJD, Upchurch P & Goswami A. 2016 Eutherians experienced elevated evolutionary rates in the immediate aftermath of the Cretaceous–Palaeogene mass extinction. Proceedings of the Royal Society B 283:20153026, 1-8. DOI: https://doi.org/10.1098/rspb.2015.3026
- Hartwig WC (ed.) 2002. The Primate Fossil Record. Cambridge University Press, Cambridge (UK), xii+530 pp.
- Hooker, J. J.. Russell, D. E., & Phélizon, A. (1999). A new family of Plesiadapiformes (Mammalia) from the Old World Lower Paleogene. Palaeontology 42(3), 377–407. DOI: https://doi.org/10.1111/1475-4983.00078
- Hoose Nv 2018. Oldest-known ancestor of modern primates may have come from North America, not Asia. Florida Museum November 29, 2018. https://www.floridamuseum.ufl.edu/science/oldest-primates-north-america/
- Kay RF, Thewissen JGM & Yoder AD 1992. Cranial anatomy of Ignacius graybullianus and the affinities of the Plesiadapiformes. American Journal of Physical Anthropology 89(4), 477–498. DOI: https://doi.org/10.1002/ajpa.1330890409
- Kemp TS 2005. The Origin and Evolution of Mammals. Oxford University Press, Oxford (UK), 331 pp. [Google Books]
- Lillegraven JA, Kielan-Jaworowska Z & Clemens WA (eds) 1979. Mesozoic Mammals The First two-thirds of Mammalian History. University of California Press, Berkeley (CA), x+311 pp.
- Lofgren DL, Lillegraven JA, Clemens WA, Gingerich PD & Williamson TE 2004. Paleocene Biochronology: The Puercan Through Clarkforkian Land Mammal Ages. Chapter 3, pp. 43–105 in: Woodburne M (ed.). Late Cretaceous and Cenozoic Mammals of North America – Biostratigraphy and Geochronology. Columbia University Press, New York (NY), 400 pp. DOI: https://doi.org/10.7312/wood13040-005
- Marivaux L, Tabuce R, Lebrun R, Ravel A, Adaci M, Mahboubi MH & Bensalah M 2011. Talar morphology of azibiids, strepsirhine-related primates from the Eocene of Algeria: Phylogenetic affinities and locomotor adaptation. Journal of Human Evolution 61(4), 447–457. DOI: https://doi.org/10.1016/j.jhevol.2011.05.013
- McKenna MC & Bell SK 1997. Classification of Mammals above the Species Level. Columbia University Press, New York (NY), xii+631 pp.
- McKeever A 2021. Did ancient primates walk alongside T. rex? New evidence backs up theory. National Geographic News March 25, 2021. https://www.nationalgeographic.com/science/article/did-ancient-primates-walk-alongside-dinosaurs-new-evidence-backs-up-theory
- Morse PE, Chester SGB, Boyer DM, Smith T, Smith R, Gigase P & Bloch JI 2019. New fossils, systematics, and biogeography of the oldest known crown primate Teilhardina from the earliest Eocene of Asia, Europe, and North America. Journal of Human Evolution 128, 103–131. DOI: https://doi.org/10.1016/j.jhevol.2018.08.005
- Ni X, Wang Y, Hu Y & Li C 2004. A euprimate skull from the early Eocene of China. Nature427(6969), 65–68. DOI: https://doi.org/10.1038/nature02126
- Ni X, Hu Y, Wang Y & Li C 2005. A clue to the Asian origin of euprimates. Anthropological Science 113(1), 3–9. DOI: https://doi.org/10.1537/ase.04s001
- Ni X, Gebo D, Dagosto M, Meng J, Tafforeau P, Flynn JJ & Beard KC 2013. The oldest known primate skeleton and early haplorhine evolution. Nature 498(7452), 60–64. DOI: https://doi.org/10.1038/nature12200
- Ni X, Li Q, Li L & Beard KC 2016. Oligocene primates from China reveal divergence between African and Asian primate evolution. Science 352(6286), 673–677. DOI: https://doi.org/10.1126/science.aaf2107
- Nickerson C 2008. A long trek for ancient mini monkeys. The Boston Globe March 3, 2008. http://archive.boston.com/news/science/articles/2008/03/03/a_long_trek_for_ancient_mini_monkeys/
- Rose KD 1994. The earliest primates. Evolutionary Anthropology 3(5), 159–173. DOI: https://doi.org/10.1002/evan.1360030505
- Rose KD 2006. The Beginning of the Age of Mammals. John Hopkins University Press, Baltimore (MA), xiv+428 pp.
- Rose KD, Godinot M & Bown TM 1994. The Early Radiation of Euprimates and the Initial Diversification of Omomyidae. pp. 1–28 in: Fleagle JG & Kay RF (eds). Anthropoid Origins. Plenum Press, New York (NY), 708 pp. DOI: https://doi.org/10.1007/978-1-4757-9197-6_1
- Rose KD, Chester SGB, Dunn RH, Boyer DM, Chew AE & Bloch JI 2011. New fossils of the oldest North American euprimate Teilhardina brandti (Omomyidae) from the Paleocene-Eocene Thermal Maximum. American Journal of Physical Anthropology 146(2), 281–305. DOI: https://doi.org/10.1002/ajpa.21579
- Sanders R 2021. Our earliest primate ancestors rapidly spread after dinosaur extinction. Berkeley News February 24, 2021. https://news.berkeley.edu/story_jump/our-earliest-primate-ancestors-rapidly-spread-after-dinosaur-extinction/
- Seiffert ER, Tejedor MF, Fleagle JG, Novo NM, Cornejo FM, Bond M, de Vries D & Campbell jr KE 2020. A parapithecid stem anthropoid of African origin in the Paleogene of South America. Science 368(6487), 194–197. DOI: https://doi.org/10.1126/science.aba1135
- Sigé B, Jaeger JJ, Sudre J & Vianey-Liaud M 1990. Altiatlasius koulchii n. gen. et sp., primate omomyidé du Paléocène supérieur du Maroc, et les origines des euprimates. Palaeontographica A 214(1-2), 31–56. https://www.schweizerbart.de/papers/pala/detail/A214/71382/
- Silcox MT 2001. A phylogenetic analysis of Plesiadapiformes and their relationship to Euprimates and other archontans. Ph.D. Dissertation, Johns Hopkins University, School of Medicine.
- Silcox MT 2014. Primate Origins and the Plesiadapiforms. Nature Education Knowledge 5(3), 1. https://www.nature.com/scitable/knowledge/library/primate-origins-and-the-plesiadapiforms-106236783/
- Silcox MT, Boyer DM, Bloch JI & Sargis EJ 2007. Revisiting the adaptive origins of primates (again). Journal of Human Evolution 53(3), 321–324. DOI: https://doi.org/10.1016/j.jhevol.2007.01.010
- Silcox MT, Bloch JI, Boyer DM, Chester SGB & López-Torres S 2017. The evolutionary radiation of plesiadapiforms. Evolutionary Anthropology 26(2), 74–94. DOI: https://doi.org/10.1002/evan.21526
- Smith T, Rose KD & Gingerich PD 2006. Rapid Asia–Europe–North America geographic dispersal of earliest Eocene primate Teilhardina during the Paleocene–Eocene Thermal Maximum. PNAS 103(30), 11223–11227. DOI: https://doi.org/10.1073/pnas.0511296103
- Soligo, C. & Martin, R.D. The first primates: a reply to Silcox et al. Journal of Human Evolution 53(3), 325-328 (2007). https://doi.org/10.1016/j.jhevol.2007.05.003
- Szalay FS & Delson E 1979. Evolutionary History of the Primates. Academic Press, London (UK), xiv+580 pp. DOI: https://doi.org/10.1016/C2009-0-22035-6
- Sudre J 1975. Un prosimien de Paleogene ancien du Sahara nord-occidental: Azibius trerkin. g. n. sp. Comptes rendus de l’Academie des Sciences de Paris D 280, 1539–1542. https://gallica.bnf.fr/ark:/12148/bpt6k57685706/f565.item
- Tabuce R, Mahboubi M, Tafforeau P & Sudre J 2004 Discovery of a highly specialized Plesiadapiformes (Mammalia, Primates) in the Eocene of Africa. Journal of Human Evolution 47(5), 305–321. DOI: https://doi.org/10.1016/j.jhevol.2004.08.005
- Tabuce R, Marivaux L, Lebrun R, Adaci M, Bensalah M, Fabre P-H, Fara E, Gomes Rodrigues H, Hautier L, Jaeger J-J, Lazzari V, Mebrouk F, Peigne S, Sudre J, Tafforeau P, Valentin X & Mahboubi M 2009. Anthropoid versus strepsirhine status of the African Eocene primates Algeripithecus and Azibius: Craniodental evidence. Proceedings of the Royal Society B 276(1676), 4087–4094. DOI: https://doi.org/10.1098/rspb.2009.1339
- Van Valen L 1994. The origin of the plesiadapid primates and the nature of Purgatorius. Evolutionary Monographs 15, 1–79.
- Van Valen LM & Sloan RE 1965. The earliest Primates. Science 150(3697), 743–745. DOI: https://doi.org/10.1126/science.150.3697.743
- Wible JR, Rougier GW, Novacek MJ & Asher RJ 2007. Cretaceous eutherians and Laurasian origin for placental mammals near the K/T boundary. Nature 447(7147), 1003–1006. DOI: https://doi.org/10.1038/nature05854
- Wible JR, Rougier GW, Novacek MJ & Asher RJ 2009. The eutherian mammal Maelestes gobiensis from the Late Cretaceous of Mongolia and the phylogeny of Cretaceous Eutheria. Bulletin of the American Museum of Natural History 327, 1–123. http://hdl.handle.net/2246/6001
- Wikipedia 2022. Evolution of primates. https://en.wikipedia.org/wiki/Evolution_of_primates
- Wikipedia 2022. List of fossil primates. https://en.wikipedia.org/wiki/List_of_fossil_primates
- Wilson Mantilla GPW, Chester SGB, Clemens WA, Moore JR, Sprain CJ, Hovatter BT, Mitchell WS, Mans WW, Mundil R & Renne RR 2021. Earliest Palaeocene purgatoriids and the initial radiation of stem primates. Royal Society Open Science 8(2):210050, 1–10. DOI: https://doi.org/10.1098/rsos.210050