 Life Sciences
Life Sciences
Aquatic Bladderworts — Michael Behe’s “Irreducibly Complex” Mousetrap in Nature
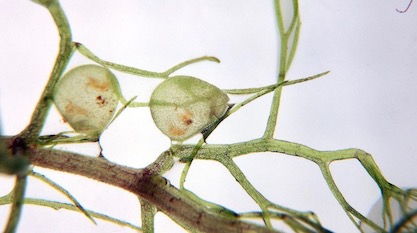
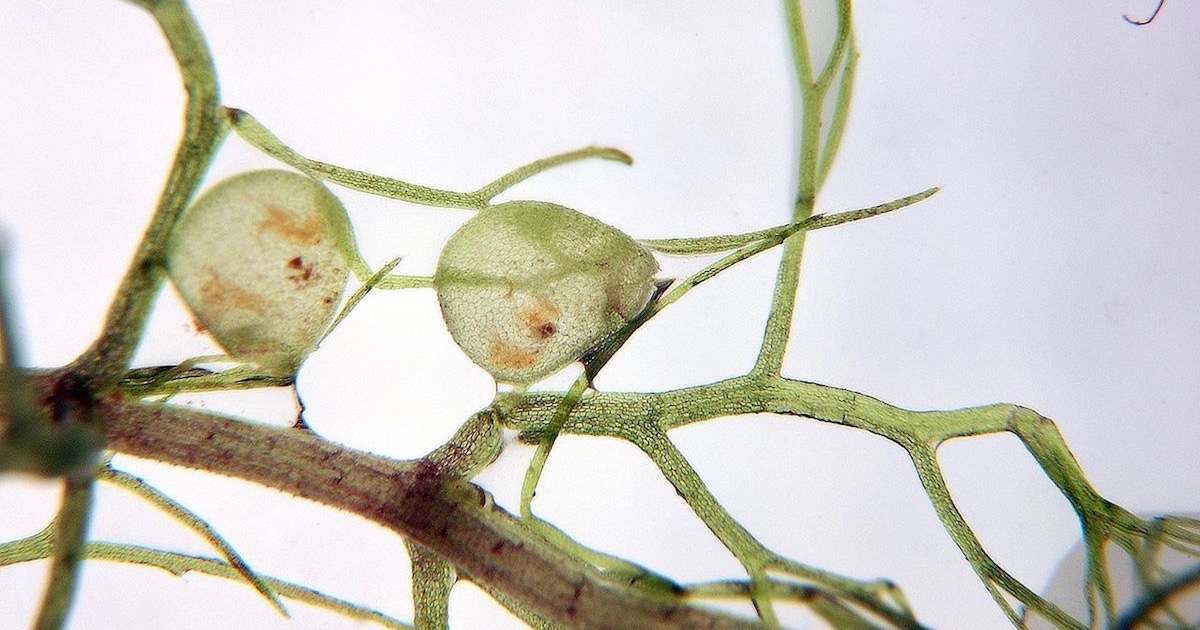
In his book Darwin’s Black Box, Michael Behe offered the mousetrap as an example of a simple everyday device that is “irreducibly complex,” because it requires several components before it can catch any mice. Behe argued that there are many such machines in living cells which have no use, and therefore provide zero selective advantage, until all or many of their numerous components are in place.
Well, here is Behe’s mousetrap in nature!
“Delicate” but Deadly
In an appendix to my 1985 book Analysis of a Finite Element Method: PDE/PROTRAN, I offered as an example of irreducible complexity (though Behe would not coin that term until 11 years later), the aquatic bladderworts (Utricularia). They are described in Plants and Environment (R.F. Daubenmire, Wiley, 1947) as follows:
The aquatic bladderworts are delicate herbs that bear bladder-like traps 5mm or less in diameter. These traps have trigger hairs attached to a valve-like door which normally keeps the trap tightly closed. The sides of the trap are compressed under tension, but when a small form of animal life touches one of the trigger hairs the valve opens, the bladder suddenly expands, and the animal is sucked into the trap. The door closes at once, and in about 20 minutes the trap is set ready for another victim.
In their article on “Carnivorous Plants” in Nature Encyclopedia of Life Sciences (Nature Publishing Group, Wiley Interscience, 2005), W.E. Loennig and Heinz-Albert Becker of the Max Planck Institute for Plant Breeding Research discuss the origin of the carnivorous plants:
Also, several authors have discussed the question whether the modern synthesis (selection of mutations “with small or even invisible effects on the phenotype”) could provide a sufficient explanation of the many convergently arisen synorganized trap mechanisms. Darwin in 1875 examined several basic points on how the origin of some carnivorous structures could perhaps be envisioned by a process of gradual evolution. However, most authors have remained critical. The following questions and statements posed by Nachtwey in 1959 concerning the origin of Utricularia’s trap still have not been answered satisfactorily at the beginning of the twenty-first century and may illustrate some of the principal problems of the origin of carnivory in plants. After a careful description of the structures and functions of the trap, Nachtwey raised the question of how the origin from a leaf tip should be envisioned and went on to ask (pp. 99-100): “Which nondirectional mutation should have occurred first in a normal leaf tip and subsequently displayed any selective advantage? Without an advantage it would have been lost as trivial. The modern synthesis strongly emphasizes that mutation and selection have to cooperate to generate new structures. So, by which blind mutations should the suction trap have originated?” And regarding the problem of further evolutionary stages the writer continues: “Even a perfect suction trap displaying the astonishing ability to rapidly catch animals would have no advantage in the struggle for life because the prey would not be digested. Conversely, the production of highly effective digestive juices would be of no avail for the tip of a leaf as long as it could not capture the prey, which is absolutely necessary. But even if suction trap and digestive juices cooperated, nothing would be gained in the struggle for life. The dissolved proteins must also be absorbed and metabolized to species-specific proteins. The formation of the suction trap requires the perfect cooperation of many different genes and developmental factors. At the end a benefit is reached in the struggle for life, but not by any evolutionary stage.” Nachtwey concluded that none of the contemporary evolutionary theories was able to answer these questions, proposing that the answer might lie outside the present scientific paradigms. Although other scenarios could possibly be envisioned and some objections raised against Nachtwey’s reflections (perhaps unknown functions of nascent structures, perhaps minor uptake of organic compounds by leaves without digestive and absorbent glands), the problem appears to have been aggravated by the recent discoveries that the well-being of Utricularia purpurea appears to be largely independent of its prey (see details under nutrition and digestion above).
Juniper et al., who favour the functional proposition of Schnell quoted above, also agree with Nachtwey on the intricacy of the evolutionary problem, stating that “no adequate evolutionary sequence can yet be constructed even to present a speculative path for the origin of what appears to be a relatively homogeneous group.” As for this problem, they continue that Utricularia “is a complex bit of mechanism and offers, as yet, an intractable problem in evolution.” The latter writers generally favour a more or less saltational model for the origin of carnivorous plants, a view shared by some other biologists as a general model for the origin of large numbers of morphological species and genera in the living world.
Yet, even authors preferring “gradual evolutionary change through unimaginable aeons of time” (Slack, 2001, p. 19) admit the depth of the origins problem for carnivorous plants: “Unfortunately this is a question which we cannot hope to answer without suitable fossil evidence, and one can offer a mere hypothesis” (Slack pp. 18-19). Moreover, it appears to be hard even to imagine clearcut selective advantages for all the thousands of postulated intermediate steps in a gradual scenario, not to mention the formulation and examination of scientific (i.e., testable) hypotheses for the origin of the complex carnivorous plant structures examined above.
The reader is further invited to consider the following problem. Charles Darwin provided a sufficiency test for his theory (1859, p. 219): “If it could be demonstrated that any complex organ existed, which could not possibly have been formed by numerous, successive, slight modifications, my theory would absolutely break down.” Darwin, however, stated that he could “not find out such a case”. Biochemist Michael J. Behe (1996, p. 39) has refined Darwin’s statement by introducing and defining his concept of “irreducibly complex systems”, specifying: “By irreducibly complex I mean a single system composed of several well-matched, interacting parts that contribute to the basic function, wherein the removal of any one of the parts causes the system to effectively cease functioning.”
Some biologists believe that the trap mechanism(s) of Utricularia and several other carnivorous plant genera (Dionaea, Aldovanda, Genlisea) come at least very near to “such a case” of irreducible complexity.
Photo: The natural “mousetrap” of Utricularia aurea, by Michal Rubeš / CC BY 3.0 CZ.
{kind=link}