 Evolution
Evolution
 Paleontology
Paleontology
Fossil Friday: The Mess of Arachnid Phylogeny, and Why I’ve Become More Skeptical of Common Descent

This Fossil Friday features the primitive spider Chimerarachne yingi from mid-Cretaceous Burmese amber (about 100 million years old). Its discovery and description by Wang et al. (2018) and Huang et al. (2018) was a major scientific sensation and celebrated as an alleged confirmation of evolutionary predictions (University of Kansas 2018). I will use this opportunity to discuss some problems of arachnid phylogeny and their implications for the assumed support of the common descent hypothesis. Anybody who has watched my lectures online or read my previous articles should know that I always emphasize that my main critique of Darwinian evolution concerns the unguided process, not the age of the Earth or common descent, which I have explicitly affirmed as the most elegant explanation of the total body of evidence. However, I have recently come to realize that the assumed evidence for common descent becomes much less convincing the closer you look into the details. I even discovered that I have to give up one of my very favorite arguments for common descent. Since this case is cumulative and somewhat complicated, please bear with me if this article gets a bit lengthy. However, I promise you will learn something important from it, even if you should be an evolutionist and skeptical of my anti-Darwinian conclusions.
What Is an Arachnid?
Arachnids are a group of land-living arthropods that many lay people find disgusting rather than interesting, even though the stunning capabilities of orb-weaving spiders certainly are fascinating. Together with the marine sea spiders (Pycnogonida or Pantopoda), marine horseshoe crabs (Xiphosura), and extinct sea scorpions (Eurypterida), the terrestrial Arachnida belong to an ancient group of arthropods called Chelicerata because of their shared chelate mouth parts that resemble a pair of scissors (Weygoldt & Paulus 1979, Shultz 1990, 2007, Dunlop 1997, 2010, Dunlop & Selden 1998, Selden & Dunlop 1998, Dunlop & Arango 2004, Paulus 2004, Lamsdell 2012, 2016, Legg et al. 2013, Dunlop et al. 2014, Lamsdell et al. 2015, Giribet 2018, Bicknell et al 2019, Giribet & Edgecombe 2019, Howard et al. 2019, 2020). Arachnids include more than 110,500 described living and fossil species that are classified in several quite different orders such as scorpions (Sorpiones), mites (Acariformes and Parasitiformes incl. Opilioacarida), pseudoscorpions (Pseudoscorpiones), harvestmen (Opiliones), micro-whip scorpions (Palpigradi), sun spiders or camel spiders (Solifugae or Solpugida), hooded tickspiders (Ricinulei), whip scorpions (Thelyphonida: Uropygi and Schizomida), whip spiders (Amblypygi), and true spiders (Araneae), as well as the four extinct Paleozoic orders Haptopoda, Phalangiotarbida, Trigonotarbida, and Uraraneida.
The phylogenetic relationships and evolutionary history of arachnids has been a highly contentious issue for many decades, and is a prime example of the failure of evolutionary biology to find a congruent pattern of nested similarities. The latter is often postulated as major evidence in favor of evolution by popularizers of Darwinism such as Richard Dawkins, who has even claimed that the reconstructed patterns of relationship from different sources of evidence are all perfectly matching (Luskin 2023). We will see that nothing could be further from the truth and that this alleged major evidence for common descent does not stand up to scrutiny.
Arachnid Monophyly and Terrestrialisation
The first and most basic problem of arachnid phylogeny is the question of whether arachnids are monophyletic at all, and if an adaptation to terrestrial life originated only once or multiple times independently within arachnids. In spite of a few early dissenters (e.g., Kraus 1976, van der Hammen 1986, 1989, Selden & Jeram 1989, Selden & Dunlop 1998), the prevailing textbook wisdom has been for decades that terrestrial arachnids constitute a monophyletic group, based on a single terrestrialization of their common ancestor (e.g., Weygoldt & Paulus 1979, Shultz 1990, 2007, Weygoldt 1998, 1999, Regier & Shultz 1998, Paulus 2004, Scholtz & Kamenz 2006, Dunlop 2010, Regier et al. 2010, Legg et al. 2013, Garwood & Dunlop 2014, Selden et al. 2015, Garwood et al. 2017, Giribet 2018, Huang et al. 2018, Wang et al. 2018, Howard et al. 2019, 2020, Lozano-Fernandez et al. 2019, 2020). Meanwhile, horseshoe crabs (Xiphosura) and the extinct eurypterids retained a marine way of life and fully developed compound eyes (Miether & Dunlop 2016, Schoenemann et al. 2019) as outgroups to Arachnida.
Ballesteros & Sharma (2019) commented that
the dominant hypothesis has been that horseshoe crabs represent the sister lineage to the terrestrial chelicerates, the highly diverse Arachnida (Snodgrass 1938; Weygoldt and Paulus 1979; Shultz 1990, 2007). In this scenario, extinct marine chelicerate groups like Eurypterida (sea scorpions) and Chasmataspidida are inferred to constitute a grade subtending Arachnida (Dunlop and Webster 1999). Implicit in this hypothesis of a monophyletic Arachnida is the notion of a single transition to the terrestrial environment by the common ancestor of arachnids. This hypothesis is supported in part by the morphological correspondence between the respiratory organs of horseshoe crabs (the book gills) and the counterparts of some arachnid groups such as spiders and scorpions (the book lungs, which resemble internalized gills; Scholtz and Kamenz, 2006; Kamenz et al., 2008).
“Molecular clock dating has suggested a Cambrian-Ordovician terrestrialization event for arachnids [Lozano-Fernandez et al. 2020], some 60 Ma before their first fossils in the Silurian” (Lamsdell et al. 2020). Such a mismatch of molecular clock datings and the actual fossil record is a well-known and ubiquitous problem in paleobiology and evolutionary biology. It is yet another piece of evidence that is unexpected under Darwinism and arguably counts as good evidence against common descent. After all, if common descent holds, different lines of evidence should converge to one true history of life.
The above-mentioned standard view of monophyletic arachnids has been questioned by more recent phylogenomic studies (see Schwager et al. 2015). Sharma et al. (2014) therefore commented that
the monophyly of Arachnida, the terrestrial chelicerates, is generally accepted, but has garnered little support from molecular data” (also see this webpage of Sharma on chelicerate phylogenomics). Similarly, Ballesteros et al. (2022) admitted that “although conflicting hypotheses prevail in morphological and molecular data sets alike, the monophyly of Arachnida is nearly universally accepted, despite historical lack of support in molecular data sets.
But the problem lies not just in conflicting molecular data, but even in the morphological adaptations to a terrestrial life, which often show a high degree of convergence. In their study of water-to-land transitions in arthropods, Dunlop et al. (2013) casually remarked that it “may seem trivial, but for the major terrestrial lineages of arthropods, there are surprisingly few unambiguous examples of anatomical terrestrial adaptations defining monophyletic groups.” Sorry, but this is certainly not trivial at all, but rather represents the polar opposite of what a Darwinian theory would predict to find.
Some experts considered an assumed marine life of Paleozoic scorpions as primitive state (Kjellesvig-Waering 1986, Selden & Jeram 1989, Selden & Dunlop 1998, Dunlop & Webster 1999), which arguably could support a sister group relationship of scorpions and all other arachnids and/or suggest an independent terrestrialisation from other arachnids (Dunlop 1997, Dunlop & Selden 1998, Paulus 2004, Lamsdell et al. 2015, Lamsdell 2016, Selden et al. 2015, Aria & Caron 2019 SI, Bicknell et al. 2019 SI). However such a marine lifestyle of early scorpions turned out to be highly contentious and mainly based on the depositional environment, while morphological evidence (especially from the book lungs) rather suggests a terrestrial adaptation (Scholtz & Kamenz 2006, Dunlop et al. 2008, 2013, 2014, Kamenz et al. 2008, Kamenz 2009, Kühl et al. 2012, Waddington et al. 2015, Howard et al. 2019). Dunlop et al. (2013, 2014) therefore found that “the trend seems to be shifting towards interpreting all fossil scorpions as potentially terrestrial animals” or “the trend is now to see most, if not all, fossil scorpions as terrestrial.”
Much Too Generous in My Assessment
Incidentally, a few years ago I wrote an article for Evolution News (Bechly 2020), in which I harshly critiqued the description of the alleged earliest scorpion Parioscorpio venator from the Lower Silurian of Wisconsin and the evolutionary speculations about arachnid terrestrialization that were boldly built upon this fossil discovery by Wendruff et al. (2020) and readily adopted by the pop science media (e.g. Neethling 2021). Just a year later a new study by Anderson et al. (2021) showed that I was even much too generous in my assessment, because these authors debunked any scorpion affinity of Parioscorpio and placed this fossil in an uncertain but much more basal position among the so-called great appendage arthropods. All the evolutionary speculations turned out to be junk science exactly as I had said. But, at that time my view was of course ignored as nothing but creationist bovine excrement. God forbid that intelligent design proponents might be correct with their critique of evolutionary speculations and are even making successful predictions.
Anyway, some experts had previously argued for a closer relationship of sea scorpions (Eurypterida) and true scorpions (e.g., Grasshoff 1978, Kjellesvig-Waering 1986, Smith 1990, Starobogatov 1990, Braddy et al. 1999, Dunlop & Webster 1999, Dunlop & Braddy 2001; also see discussion in Dunlop 1997), which would of course also question arachnid monophyly and a single terrestrialization. However, their views have been criticized by Shultz (1990) as based mainly on overall similarity instead of specific similarities (synapomorphies). Today a closer relationship of sea scorpions and scorpions is mostly obsolete and has few if any supporters.
Other experts made cladistic or phylogenomic analyses and recovered scorpions in a subordinated position within Arachnids. The majority of these studies suggested a sister group relationship of scorpions and Pedipalpi+Araneae within a clade Arachnopulmonata (Regier et al. 2010, Sharma et al. 2014, Giribet 2018, Starrett et al. 2016, Ballesteros et al. 2019, Ballesteros & Sharma 2019, Giribet & Edgecombe 2019, Howard et al. 2019, Lozano-Fernandez et al. 2019, 2020, Noah et al. 2020, Anderson et al. 2021), which share the same type of book lungs. However, other scientists disagreed and instead suggested scorpions as sister group of pseudoscorpions (Pepato et al. 2010, Dunlop et al. 2014, Garwood & Dunlop 2014, Garwood et al. 2016, 2017, Huang et al. 2018, Wang et al. 2018, Howard et al. 2020, Ontano et al. 2021, Ballesteros et al. 2022, Dunlop 2022), or as sister group of pseudoscorpions and camel spiders (Shultz 1989, 1990, Selden & Dunlop 1998, Wheeler & Hayashi 1998, Pollitt et al. 2004), or as sister group of harvestmen (Shultz 2007, Dunlop 2010, Legg et al. 2013, Wolfe 2017 SI, Ban et al. 2022), all of which lack book lungs. Some of the studies that proposed a clade of scorpions and pseudoscorpions still placed this clade within Arachnopulmonata, which would imply that pseudoscorpions secondarily lost any trace of the defining book lungs, in spite of retaining a terrestrial way of life, which hardly makes any sense.
The recent discovery of air-breathing structures in an exceptionally preserved 340-million-year-old sea scorpion (Lamsdell et al. 2020), as well as the discovery “that respiratory organs of a new fossil eurypterid resemble arachnid book lungs, [are] supporting the hypothesis that eurypterids — and perhaps arachnid ancestors — were amphibious” (Dunlop 2020). Since the latter author recognized that “modern phylogenetic analyses do not support the hypothesis that eurypterids are specifically the closest relatives of scorpions,” the new finding could also call into question the status of book lungs as a putative synapomorphy of Arachnopulmonata (scorpions, whip scorpions, whip spiders, and spiders) that was suggested by Sharma et al (2014) and has been increasingly supported by modern studies (see Dunlop 2022).
Lamsdell et al. (2020) readily admitted that a considerable number of independent (convergent) losses and gains of complex characters would be implied either way, if Arachnida is monophyletic or polyphyletic (Lamsdell et al. 2020: fig. 3). They commented that
eurypterids were experimenting with modes of terrestrial respiration and were in the process of terrestrializing rather than returning to aquatic environments. This in turn suggests that horseshoe crabs evolved from fully aquatic ancestors. Assuming a single terrestrialization event for Arachnida therefore necessitates that non-pulmonate arachnids lost their book lungs (Figure 3B), with tracheae evolving multiple times among non-pulmonates, as indicated by their occurrence on different body segments in different groups [26]. Alternatively, arachnids may have invaded land multiple times [42]; however, this scenario still necessitates that non-pulmonates lost their respiratory lamellae and independently developed tracheae.
If you find this sounds confusing and wishy-washy, lacking any scientific rigor, you are certainly not alone. This is the typical “anything goes” just-so storytelling that dominates modern evolutionary biology and makes it such a frustrating discipline.
But It Gets Even Weirder
Van der Hammen (1986, 1989) recovered the aquatic horseshoe crabs and scorpions as sister group of harvestmen deeply nested within terrestrial arachnids. This fringe view was largely forgotten until recently, when Sharma et al. (2014, 2021), Ballesteros et al. (2019), and Ballesteros & Sharma (2019) recovered horseshoe crabs within Arachnida as sister group to the small order Ricinulei (also see Ontano et al. 2021). This was supported by a more comprehensive study by Ballesteros et al. (2022), which even suggested the “resurrection” of a marine clade Merostomata (Xiphosura, Synziphosurina, Chasmataspidida, and Eurypterida) in this subordinate position, contrary to the conventional modern view of these fossil groups as basal marine grade in the stem group of Arachnida (e.g., Cotton & Braddy 2004, Lamsdell et al. 2015, Lamsdell 2016, Selden et al. 2015). This would not just imply a non-monophyly of Arachnida and a multiple independent terrestrialization, but would also imply a convergence of the very similar breathing organs (book lungs) and the convergent reduction of the compound eyes, which appeared to be congruent with assumed aquatic Paleozoic scorpions with compound eyes (Selden & Jeram 1989, Selden & Dunlop 1998, Miether & Dunlop 2016, Schoenemann et al. 2019). The authors therefore concluded that:
Combined analyses recovered the clade Merostomata (the marine orders Xiphosura, Eurypterida, and Chasmataspidida), but merostomates appeared nested within Arachnida. Our results suggest that morphological convergence resulting from adaptations to life in terrestrial habitats has driven the historical perception of arachnid monophyly, paralleling the history of numerous other invertebrate terrestrial groups.
This study was by no means the work of a fringe group of maverick cranks, but included some of the most distinguished experts of arthropod phylogenetics as co-authors, such as Gonzalo Giribet, Mark Harvey, Prashant Sharma, and Ward Wheeler. Another recent cladistic analysis of mitochondrial genomes by Ban et al. (2022) came to a similar result, with horseshoe crabs subordinated within arachnids as sister group of camel spiders, while a supermatrix analysis of Noah et al. (2020) placed horseshoe crabs as sister group of a clade Scorpiones+Pedipalpi+Araneae. However, the latter authors differed from all others in suggesting that “the ancestor of Xiphosura and the extinct Eurypterida (sea scorpions, of which many later forms lived in brackish or freshwater) returned to the sea after the initial chelicerate invasion of land.” Obviously, nothing is forbidden in evolutionary fantasy land as long as the fundamental paradigm of common descent with unguided modification is not questioned.
Let this really sink in: According to the most recent and most comprehensive studies, the previous decades of phylogenetic trees, evolutionary scenarios, and reconstructed ancestors (ground plans) would all be utterly incorrect. Alternatively, if the most modern studies are wrong (as for example implied by the recent cladistic study of fossil evidence by Anderson et al. 2021, as well as phylogenomic studies of Lozano-Fernandez et al. 2019, 2020 and Shingate et al. 2020), then even the most advanced methodologies and most comprehensive data sets would lead to incorrect evolutionary hypotheses. Either way, you cannot ignore that evolutionary biology is a state of disarray. Something is clearly and profoundly off the mark and conflicting with any expectations from Darwinian theory. I can only urge my colleagues to stop closing their eyes, only because of world view blinders, and recognize the obvious need for a paradigm change, because we have just scratched the surface of the problems. There is more — much more.
The Enigma of Arachnid Interordinal Relationships
Also, the phylogenetic reconstructions of the interrelationships of the different arachnid orders are a total mess as different studies (e.g., Weygoldt & Paulus 1979, van der Hammen 1986, 1989, Schultz 1989, 1990, 2007, Smith 1990, Selden & Dunlop 1998, Weygoldt 1998, 1999, Wheeler & Hayashi 1998, Dunlop & Webster 1999, Giribet et al. 2002, Harvey 2002, Paulus 2004, Pollitt et al. 2004, Dunlop 2010, 2022, Pepato et al. 2010, Regier et al. 2010, Legg et al. 2013, Dunlop et al. 2014, Garwood & Dunlop 2014, Sharma et al. 2014, 2021, Lamsdell et al. 2015, Schwager et al. 2015, Selden et al. 2015, Garwood et al. 2016, 2017, Lamsdell 2016, Starrett et al. 2016, Wolfe 2017, Giribet 2018, Huang et al. 2018, Wang et al. 2018, Aria & Caron 2019, Ballesteros & Sharma 2019, Ballesteros et al. 2019, 2022, Bicknell et al 2019, Giribet & Edgecombe 2019, Howard et al. 2019, 2020, Lozano-Fernandez et al. 2019, 2020, Noah et al. 2020, Anderson et al. 2021, Ontano et al. 2021, 2022, Ban et al. 2022) produced very different trees with hardly any agreement on specific groupings (see Wheeler & Hayashi 1998, Giribet 2018 and Sharma). There is no wonderful consensus on a single tree of life, and people like Richard Dawkins are either utterly clueless or deliberately lying when they claim otherwise.
Actually, none of the suggested phylogenies can plausibly accommodate the very incongruent distribution of highly complex similarities in the circulatory system described by Göpel & Wirkner (2015: fig. 12) for horseshoe crabs and some arachnids. It is not just that morphological versus molecular data produce conflicting trees, or that different phylogenetic methods produce different trees, but even cladistic studies of traditional anatomy produced totally different trees (e.g., Weygoldt & Paulus 1979 vs Shultz 1990) that could not be resolved to this day. A strict consensus tree (think of a kind of lowest common denominator) of all the published phylogenies of arachnids would indeed result in an unresolved polytomy or lawn instead of a branching tree (Sharma et al. 2021: fig. 1). This is literally a collapse of phylogenetic theory and its predictions from common descent with modification.
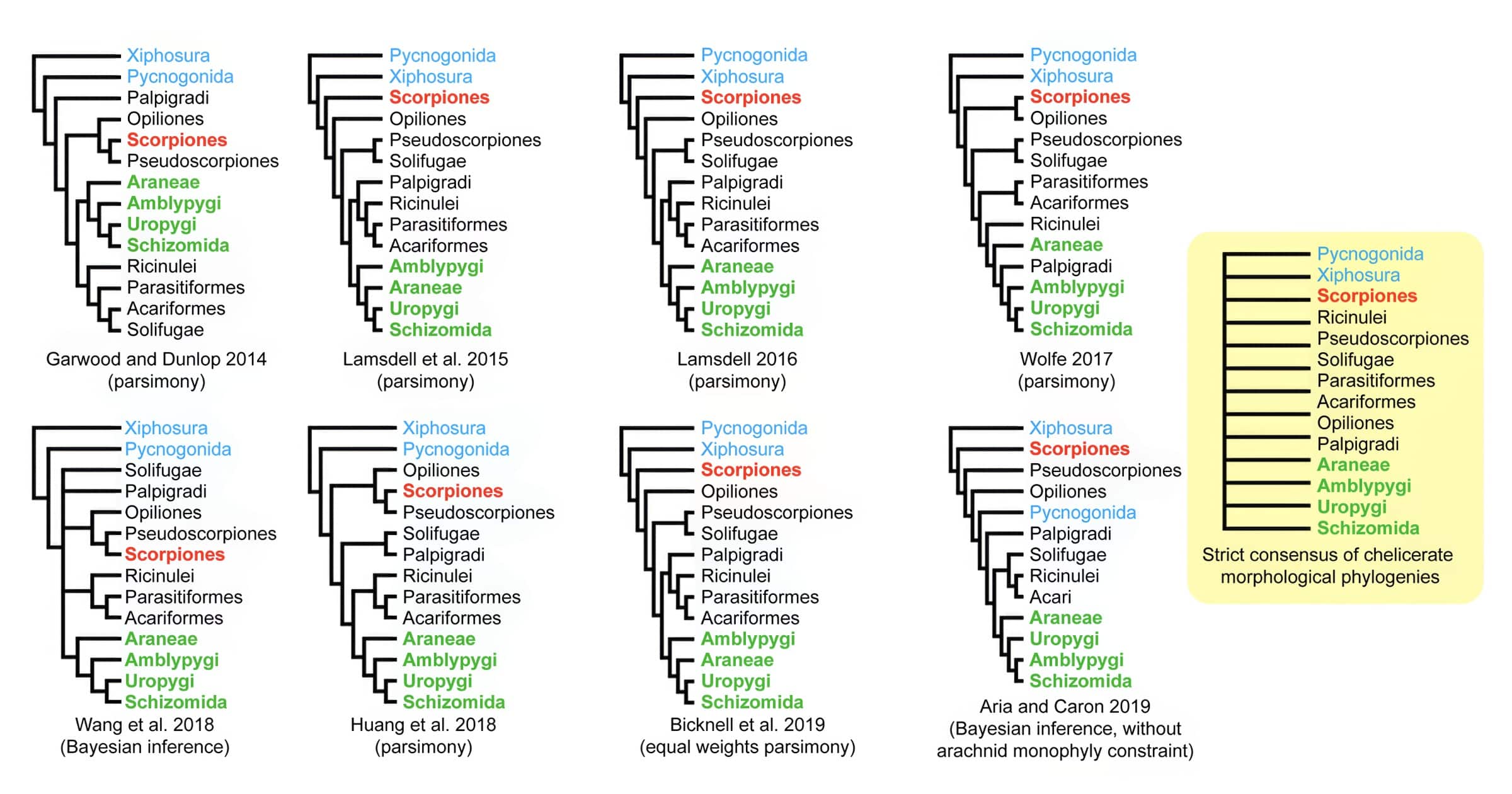
Therefore, Sharma et al. (2014) admitted: “Attempts to resolve the internal phylogeny of chelicerates have achieved little consensus, due to marked discord in both morphological and molecular hypotheses of chelicerate phylogeny.” Sharma et al. (2021) commented that “the basal phylogeny of Chelicerata is one of the opaquest parts of the animal Tree of Life, defying resolution despite application of thousands of loci and millions of sites” and called this a “gordian knot in metazoan phylogeny.” Strikingly, Kuntner (2022) did not even bother to mention this crucial issue in his list of seven grand challenges in arachnid science, which is quite telling about the deplorable state of denial in evolutionary biology.
The uncertainty of the interordinal relationships of arachnids also holds for the putative closest relative (sister group) of spiders, which greatly influences all evolutionary speculations. Some earlier studies based on classical comparative morphology favoured whip spiders as closest relatives of spiders among living arachnids (Weygoldt & Paulus 1979, van der Hammen 1986, 1989, Wheeler & Hayashi 1998, Alberti & Michalik 2004, Paulus 2004), while a few outlier studies even suggested whip scorpions as sister group of spiders (Giribet et al. 2002, Pepato et al. 2010, Lamsdell et al. 2015 SI, Bicknell et al. 2019 SI). The majority of studies supported a clade (Pedipalpi) of whip scorpions plus whip spiders as sister group of spiders (Shultz 1989, 1990, 1999, 2007, Smith 1990, Selden et al. 1991, 2015, Selden & Dunlop 1998, Giribet et al. 2002, Pollitt et al. 2004, Dunlop 2010, 2022, Regier et al. 2010, Legg et al. 2013, Dunlop et al. 2014, Garwood & Dunlop 2014, Sharma et al. 2014, 2021, Garwood et al. 2016, 2017, Lamsdell 2016 SI, Starrett et al. 2016, Wheeler et al. 2016, Giribet 2018, Huang et al. 2018, Wang et al. 2018, Aria & Caron 2019 SI, Ballesteros & Sharma 2019, Ballesteros et al. 2019, 2022, Giribet & Edgecombe 2019, Howard et al. 2019, 2020, Lozano-Fernandez et al. 2019, 2020, Noah et al. 2020, Ontano et al. 2021, Ban et al. 2022). Of course, the phylogenetic framework makes a big difference for the ground plan reconstruction of spiders. We will come back to this point when we discuss the alleged confirmation of a predicted tail filament (flagellum) in primitive spiders.
The Origin of Spider Silk and Spinnerets
Another much-discussed problem for the evolution of spiders is the origin of spider silk and the spinnerets that are unique to spiders and a genuine engineering marvel (Vollrath & Selden 2007, Hilbrant 2008, Brunetta & Craig 2010, Mariano-Martins et al. 2020; also see Wikipedia).
The oldest uncontroversial spiders with preserved spinnerets belong to the genus Arthrolycosa of Carboniferous and Permian age (Selden & Penney 2010, Selden et al. 2014), while all supposed Devonian spiders were later shown to be based on misidentifications (Selden 2021).
Shear et al. (1989) had described a 380-million-year-old assumed isolated spider spinneret from the Devonian of New York, but this was shown by Selden et al. (2008) to be a spigot of Attercopus that was described as a spider by Selden et al (1991) from the same locality, but later assigned by Selden et al. (2008) to a separate Paleozoic order Uraraneida, only including the genera Attercopus and Permarachne. A homology of the silk spigots of Uraraneida and the spinnerets of spiders was suggested by Selden et al. (2008) and supported by several subsequent studies such as Legg et al. (2013), Garwood & Dunlop (2014), and Dunlop (2022). It was because of these silk glands, that Attercopus was long accepted to be the oldest and most primitive spider (Vollrath & Selden 2007), which is still often repeated in popular sciences articles (e.g., Gray 2018). Wunderlich (2015) even included Uraraneida as a suborder in the spider order Araneae, but today Uraraneida is usually considered to be the fossil sister group of spiders, even more closely related to them than are the living whip scorpions and whip spiders.
Even though Uraraneida did indeed possess silk glands, these were definitely not arranged on spinnerets (Selden et al. 2008, Selden 2021), so that the postulated homology may be questionable. But it gets worse: uraraneids also did not possess any leg-like appendages at the place where spinnerets would be, even though spider spinnerets are segmented and believed to be modified leg appendages homologous to horseshoe crab gills (Damen et al. 2002, Selden et al. 2008, Wang et al. 2018) or walking legs (Hilbrant 2008, Pechmann & Prpic 2009, Shoemaker et al. 2017). The structure from which spinnerets are thought to have evolved is totally lacking in stem spiders that clearly should possess them as precursor organs. This is definitely not at all what Charles Darwin would have predicted.
Wang et al. (2018) recognized this problem and commented:
A previous study discussed a paradox in that spider spinnerets are modified opisthosomal appendages, probably representing the original telopod. Outgroup comparison implies that retaining these limbs should be plesiomorphic, but spider relatives such as uraraneids lack opisthosomal appendages. The authors postulated that there may be a genetic mechanism in spiders that reactivated the development of (lost) appendages, allowing the evolution of movable spinnerets that facilitate a more precise manipulation of silk strands.
I discussed the problem of such assumed genetic reactivation in a recent article on insect metamorphosis (Bechly 2023b). Spider spinnerets represent yet another case, where the evo-devo and genomic evidence are ambiguous and do not converge to one true evolutionary scenario and homology hypothesis, as should be expected if common descent is true.
Another problem is that Chimerarachne from mid-Cretaceous Burmese amber, which represents the supposed primitive sister group of all spiders, does not show the predicted ancestral pattern of spinnerets. Wang (2018) admitted:
Based on the anatomy of mesotheles, we would have expected a spider ancestor to have had four pairs of spinnerets, all positioned in the middle of the underside of the abdomen. Chimerarachne only has two pairs of well-developed spinnerets, towards the back of the abdomen, with another pair apparently in the process of formation.
More failed predictions. See any pattern yet?
The uncertainty of spinneret evolution also extends to the origin of orb weaving in spiders, as was shown in a study by Bond et al. (2014), who rejected the “prevailing paradigm for orb web evolution” (also see Penney & Ortuño 2006). The whole of evolutionary biology turns out to be a house of cards — wild speculations built upon further speculations with very weak and highly ambiguous circumstantial evidence. To sell any of these evolutionary speculations as scientifically established facts, and every rewriting of previous textbook wisdom as normal scientific progress, is nothing but a great deception of a gullible public, that is mostly ignorant of the true mess behind such bold scientific claims. The truth is that the emperor has no clothes.
Like many other transitions in the history of life, the origin of spider silk and spinnerets involves a de novo origin and/or re-engineering of these complex structures, which certainly required numerous codependent mutations. This implies a significant waiting time problem to accommodate the required genetic changes in the geologically available short window of time. I have discussed this fatal problem for neo-Darwinism in several previous articles.
Tailed Spiders — A Successful Prediction of Common Descent?
When asked why, in spite of my critique of neo-Darwinism and my endorsement of intelligent design, I still subscribe to common descent, I have often answered as follows: the hallmark of a good theory are very specific predictions that are unique to this theory and successfully confirmed by later discovery of empirical evidence. The hypothesis of common descent arguably allows for the prediction of very precise anatomical details of hypothetical transitional forms, that would not be predicted by the hypothesis of common design without the constraint of shared ancestry. Such predictions have been made in the published technical literature and have indeed been confirmed by later discovered fossils of such transitional forms with precisely the predicted anatomy, which was unknown to occur either in living or fossil representatives at the time of the prediction. This even happened in my own paleoentomological work on the origin of the secondary male genital apparatus in damselflies and dragonflies. However, since this personal example is quite esoteric and requires some elaborate explanation, I have usually referred instead to the much simpler example of tailed spiders (e.g., Bechly 2021, Bechly 2023a).
This virtually convincing story goes like this: Living and fossil spiders have paired spinnerets but no median tail filament (flagellum) on their hind body. Based on the reconstructed phylogenetic relationship and the character distribution of a tail filament in living and fossil arachnids, scientists predicted the existence of primitive spiders with a tail filament and spinnerets, even though no animals with such a combination of characters were known at the time of the prediction. Then, years later primitive fossil spiders with precisely the predicted combination of spinnerets and a median tail filament were discovered in 100-million-year-old Burmese amber.
Indeed, in 2018 the sensational fossil discovery was described from mid-Cretaceous Burmese amber (Wang et al. 2018 and Huang et al. 2018; also see Wang 2018 and University of Kansas 2018). It was named Chimerarachne yingi, which shares with spiders the presence of spinnerets and of male pedipalps modified into sperm-transfer organs, but shares with uropygids and extinct uraraneids the presence of a tail filament (flagelliform telson). Apart from the latter trait, Chimerarachne could just be considered as a spider (Selden 2021). Consequently, Wunderlich (2019, 2022) had included Chimerarchne as distinct suborder Chimerarachnida within Araneida (accepted by Selden 2021 and Dunlop 2022) and described a second genus and species Parachimerarachne longiflagellum from Burmese amber. However, among the original describers there was considerable disagreement about the correct placement of the fossils, with Wang et al. (2018) including Chimerarachne among true spiders as earliest branch of the order Araneae, while Huang et al. (2018) considered it as a member of the extinct order Uraraneida (more closely related to Attercopus and Permarachne than to modern spiders). So, if Chimerarachne is a spider or not “all depends which palaeontologist you ask” (Economist 2018).
Anyway, a careful look at the actual evidence suggests that the above-mentioned successful prediction may not be as good as it sounds. To see why, let us first check if there really existed any precise prediction in the first place. Wang et al. (2018) wrote that “C. yingi preserves only part of the predicted ground pattern for spiders” but gave no reference where such a specific prediction was ever made. The press release from the University of Kansas (2018) claimed that the discovery confirmed a prediction made by Selden et al. (2008), when they described the Paleozoic order Uraraneida. However, such a specific prediction is nowhere found in this paper, but only a general statement that “the multisegmented flagellum may be a plesiomorphy of Pantetrapulmonata (13) that has been retained in Uropygi” and that:
The external mold of the London specimen of Palaeothele shows an anal tubercle, and to ascertain whether this continued into a flagellum (which could place Palaeothele as an intermediate between Araneae and the order), an X-ray computed tomography (CT) scan was performed on the specimen by M.D.S. (Fig. 3D). This showed without doubt that there is no flagellum, and therefore Palaeothele remains the earliest and only described fossil mesothele spider to date.
The authors even admitted that “It is possible that the flagellum was uniquely derived and not homologous with that of the pedipalp orders.” In other words: they “predicted” that whatever would be found would fit the evolutionary narrative. Not exactly a specific prediction at all.
So Much for That Myth
Anyway, even if the prediction was not explicitly made, was it maybe implicitly made, thus necessarily implied by the evidence? This would be the case if the most parsimonious interpretation of the evidence, based on the distribution of similarities and the reconstructed phylogenetic relationships, would place the existence of a tail filament in the ground plan of spiders. Let’s look if this is the case.
We first have to study the distribution of the crucial character of the flagellum in the assumed monophylum Tetrapulmonata, which includes whip scorpions, whip spiders, spiders, and their fossil relatives (see the tree of Tetrapulmonata in Wikipedia for a good visualization of the incongruent distribution of a tail filament in the closer spider relationship).
The only arachnids with a tail filament or flagellum are the living micro-whip scorpions (Palpigradi) and whip scorpions (Thelyphonida), as well as the extinct Uraraneida. They are neither most closely related to each other nor are they forming a basal grade of primitive arachnids. All other arachnids lack a tail filament, including whip spiders (Amblypygi) and true spiders (Araneae). Therefore, a tail filament cannot be reasonably attributed to the ground plan of arachnids, and indisputably has an incongruent distribution that suggest multiple independent gains and/or losses of the character.
So, where did the tail filament come from? The still marine horseshoe crabs, extinct chasmataspidids (and a few isolated horseshoe-crab-like families like Bunodidae and Pseudoniscidae, see Lamsdell 2012 and Selden et al. 2015), as well as Paleozoic sea scorpions (eurypterids), have a segmented hind body (metasoma) ending in a spine or sting. This terminal unsegmented element of the hind body is called telson (Snodgrass 1938; also see Lauterbach 1980) and has been universally homologized with the multisegmented tail filament, in spite of lacking any complex or specific similarity. Among terrestrial arachnids a fully developed segmented hind body with a terminal sting is only found in true scorpions (in whip scorpions the metasoma is reduced to only three short segments), but while early morphological studies recovered scorpions as the most basal branch of arachnids (Weygoldt & Paulus 1979), most modern studies of morphological and genetic data resolved scorpions in various deeply subordinated positions within arachnids (Wheeler & Hayashi 1998, Shultz 2007, Garwood & Dunlop 2014, Giribet et al. 2002, Regier et al. 2010, Sharma et al. 2014). This arguably suggests that a segmented hind body was independently reduced and the telson independently transformed into a tail filament multiple times within arachnids, so that no clear prediction can be made for the ground plan of spiders within the evolutionary paradigm. Indeed, a multiple convergent origin of a flagellum is also implied by the discovery of a Devonian sea spider with a multisegmented, flagelliform telson (Poschmann & Dunlop 2006), which belongs to marine pantopods and thus lies clearly outside of arachnids and their close relationship.
Even the oldest whip spiders from the Carboniferous lack any tail filament (Garwood et al. 2017, Dunlop 2018), and such a flagellum is also absent in the assumed fossil sister group Haptopoda of whip scorpions and whip spiders (Garwood & Dunlop 2014), which again suggests that the flagellum of whip scorpions is an autapomorphic convergence.
The fact that not just Haptopoda, but also other Paleozoic arachnid orders such as Phalangiotarbida and Trigonotarbida, which are considered as closely related to Uraraneida+Araneae and/or Pedipalpi within Tetrapulmonata, but lack any flagellum as well as any spinnerets (see Garwood & Dunlop 2014, Garwood et al. 2017, Huang et al. 2018, Wang et al. 2018, Dunlop 2022; also see Howard et al. 2019: fig. 1), further complicates the picture and even more strongly suggests a convergence.
As I have already mentioned, a tail filament has been documented for the uraraneid genera Attercopus and Permarachne by Selden et al. (2008). It is worth mentioning that “a flagellar structure was described in Permarachne (11), but because such a structure was previously unknown in spiders, yet all other morphological features suggested that Permarachne was a mesothele, the structure was interpreted as an elongate, multiarticled spinneret” (Selden et al. 2008). This revealing admission shows how much the interpretation of fossil anatomy is based on evolutionary bias and preconceived ideas. Ancient anatomy is often more hypothesis than data. But anyway, for our present purposes we can definitely code uraraneids as possessing a flagellum.
Garwood et al. (2016) described the genus Idmonarachne from the Late Carboniferous of Montceau-les-Mines in France, which is superficially similar to Uraraneida and shares their lack of spinnerets, but was proposed to be closer related to true spiders with whom it shares the lack of a flagellum as well as a similar leg segmentation and forward directed cheliceres (also see Pappas 2016). Nevertheless, other studies disagreed and recovered Idmonarachne as more distantly related to spiders than Uraraneida (Huang et al. 2018, Ballesteros et al. 2022), also because it shares divided opistosomal tergites with the extinct arachnid order Trigonotarbida (Wang et al. 2018). The incongruent pattern of similarities again and again disagrees with Darwinian expectations.
From the same Late Carboniferous locality in France the oldest known spider was described as Palaeothele montceauensis, which has no spinnerets preserved and clearly lacked a tail filament as well (Selden 1996, Selden et al. 2008).
This summer a 310–315-million-year-old fossil spider was described from the Carboniferous Piesberg locality in Germany by my colleague Jason Dunlop (2023). He is one of the leading experts on arachnid evolution, with whom I collaborated on fossil arachnids until my career-killing “coming out” as an ID proponent (Delclòs et al. 2008, Dunlop & Bechly 2015, Dunlop et al. 2015a). The fossil was named Arthrolycosa wolterbeeki and represents the oldest known true spider from Germany (see Funnell 2023). It has spinnerets but also lacks a tail filament, just like modern spiders.
For the sake of argument we will accept the current majority consensus view of the phylogeny of Tetrapulmonata. If we combine this phylogeny with the above described character distribution we arrive at the following picture: The tail filament has a highly incongruent distribution even within Tetrapulmonata. One sister group (Pedipalpi) includes a subclade (whip scorpions) with and one subclade (whip spiders) without a flagellum, while the other sister group would likewise include one subclade (Uraraneida) with and one subclade (Araneae) without flagellum. Haptopoda, the fossil sister group of Pedipalpi also lacks a flagellum. Idmonarachne, which is either the sister group of Uraraneida+Aranaeae or less likely the sister group of spiders also lacks a flagellum. Finally, all the earliest fossil spiders lack a flagellum. Given the otherwise homoplastic pattern of the occurrence of a tail filament, the hypothesis of a flagellum in the ground plan of Araneae would not be more parsimonious than a convergent origin of the flagellum in Thelyphonida and Uraraneida. Only after discovery of Chimerarachne, and if we follow its placement closer to Araneae than to Uraraneida (contra Huang et al. 2018), could we use parsimony to interpret the flagellum as plesiomorphy in the ground plan of Araneae. However, this interpretation would be a postdiction (retrodiction), not a prediction, let alone a successful prediction.
Given the above mentioned phylogenetic relationships and pattern of character distribution, and using the principle of parsimony (basically Ockham’s razor) to minimize the number of evolutionary steps (gains and losses) to optimally explain the character distribution on the given tree, we can now conclude that even within the evolutionary paradigm and applying the phylogenetic reasoning of mainstream cladistic methodology, a tail filament could not have been predicted for the ground plan of spiders. It is not just that the prediction was never made explicitly in practice, but it is not even implied by the data. No clear prediction would even have been possible based on these incongruent data, so that it would have been an unsupported and unreasonable prediction.
Consequently, we are faced with two major problems with the above-mentioned prediction:
- There is a clear temporal paradox, because all the earliest true spiders as well as all modern spiders are lacking a tail filament, while only a single taxon of mid-Cretaceous Burmese amber spider featured such a tail. This represents a very poor stratigraphic fit, which implies a remarkably long ghost lineage between Permian Uraraneida and Cretaceous Chimerarachnida. The term stratigraphic fit refers to the agreement (or lack thereof) between the stratigraphic orders of appearance of certain taxa and features with the predicted phylogenetic order of appearance (see below).
- The fact that neither the earliest true spiders nor some of the closest fossil relatives of spiders do possess a tail filament, makes the whole prediction of a tail filament in the ground plan of spiders highly questionable and actually refutes such a hypothesis as unparsimonious.
Maybe the tailed spider Chimerarachne from Burmese amber rather represents a highly derived reversal or convergence, than a preserved primitive state. In short: The alleged successful prediction turns out to be a mirage and a fluke, similar to the case of Tiktaalik (McLatchie 2012), where a confirmed prediction was later revealed to have been successful for the wrong reasons. I have to admit I fell for this erroneous example of tailed spiders, because I did not look deeply enough into the details to recognize the charade.
For this reason, I will no longer use this example without an explicit disclaimer that the case for common descent is not really strengthened by it at all. Of course, there may be other more solid examples of successful predictions from common descent, but they all have to be carefully evaluated and checked to determine if they are really supported by the evidence and do not represent other retrodictions parading as successful predictions. Also, the rare positive examples have to be weighed against the numerous failed examples, where such predictions have been decisively refuted by new fossil evidence, which is commonly indicated by media reports titled “New Fossil Discovery Rewrites the Story of [Fill in the Blank] Evolution.” Sounds familiar? Of course it does, as we regularly report about the latest rewritings at Evolution News. Given a plethora of failed predictions, a few successes are hardly surprising or noteworthy. As a German saying goes, “Even a blind squirrel can find a nut once in a while.”
Poor Stratigraphic Fit
Selden (1990) suggested that “a cladogram reflecting evolutionary events should concur with a complete fossil record in the sequence of events”. What he meant by this is the common sense view that stratigraphic order of appearance should more or less correspond to the phylogenetic order of branching in the reconstructed trees. This is called stratigraphic fit or stratigraphic congruence, and is a well-recognized concept in paleobiology (Norell & Novacek 1992, Clyde & Fisher 1997). However, stratigraphic fit is often poor, contrary to the expectation of Darwinian evolution. Unsurprisingly, this is also the case with fossil arachnids. Therefore, Shultz (1994) discussed this poor fit and simply dismissed “stratigraphic tests of phylogeny as unworkable, as they rest upon the questionable assumption that the origin of extant lineages and the origin of their diagnostic characters are coupled”. What a convenient (and unscientific) way to get rid of conflicting evidence that could refute your hypothesis! Moreover, Shultz was simply wrong, as demonstrated by botanist Armen Takhtajan’s principle of the heterobathmy of characters, which was popularized by Willi Hennig, the founder of modern phylogenetics. This principle of heterobathmy simply means that the full set of derived characters of a living group originated successively in its stem lineage, so that the only requirement is that early stem group representatives at least have to possess a single of the diagnostic characters to be identifiable as member of the group.
Therefore, in most taxonomic groups stratigraphic fit is still commonly used, and when fit is good it is of course cherished as strong support for the evolutionary hypothesis and thus common descent. However, if cases of good stratigraphic fit (Benton & Hitchin 1997) count as evidence in favor of common descent, then the many cases of poor stratigraphic fit must be counted as valid conflicting evidence, instead of being ignored or explained away with convenient ad hoc hypotheses like ghost lineages.
Misidentified Fossils
Since the seminal work of Pocock (1911), fossil arachnids from Late Carboniferous of England were believed to be the oldest araneomorph spiders and even had been classified in a distinct spider family Archaeometidae. A few years ago, these fossils were re-examined with micro-CT and turned out to be incorrectly identified, so that they had to be reinterpreted as harvestmen (Selden et al. 2016). For more than a century of modern arachnology the consensus view about the early origin of spiders was way off the mark. What else might be way off?
Well, another curious footnote is the fact that the presumed giant Carboniferous spider Megarachne, which was described by an Argentinian paleontologist (Hünicken 1980) and famously featured in the BBC documentary Walking with Monsters (2005), turned out to be nothing but the misidentified remains of a sea scorpion (Selden et al. 2005, Switek 2010). The same happened again with another supposed giant spider from the Cretaceous of China, which was described as Mongolarachne chaoyangensis by Cheng et al. (2019), but shortly after revealed with fluorescence microscopy to be a forgery that used a fossil crayfish as core (Selden et al. 2019, also see Starr 2019 and University of Kansas 2019). Embarrassing errors like these are rampant in paleontology, which would be unthinkable in hard sciences like physics. Of course, this is not by itself evidence against common descent, but rather a reminder to take any bold claims of paleontological support for common descent, such as “indisputable transitional fossils,” with a considerable grain of salt.
The Enigma of Spider Webs in Amber
Last but not least, here is another enigma, which has really bugged me as professional amber expert for a long time: In Cretaceous and Tertiary amber you can regularly find well-preserved spider webs of crisscrossed and tightly spanned threads (Saint Martin et al. 2014), sometimes even with glue droplets (Zschokke 2003, 2004, Peñalver et al. 2006, Brasier et al. 2009). Thus, it is not just plane orb-weaved webs attached to a flat inner amber surface (Schlaube), but complex 3D-networks of silk threads preserved within the amber matrix. How could such delicate structures have been preserved in sticky tree resin? Imagine you were to pore honey over such a spider web. It would of course immediately get crumbled and torn. The same applies to the dense hairs of bees and other hairy insects that are preserved upright and fluffy in amber. Nobody has ever documented experimentally how this could happen, even with the lowest viscosity tree resin known to science. Maybe some non-uniformitarian and non-actualistic processes were at work, which have yet to be identified. To be clear: I am not suggesting any miraculous stuff going on, but just want to highlight how much we simply do not know even about simple phenomena in the past.
Follow the Evidence
This should make scientists a bit humbler when they boldly propose alleged solutions to the bigger enigmas in the history of life, which will always remain highly speculative unless someone invents a time machine. We were not there to watch what happened and simply don’t know, so that every reconstruction of past events is a hypothetical inference to the best explanation based on circumstantial evidence and a lot of theoretical guesswork based on shaky assumptions. To claim that we more or less know the evolutionary history of life on Earth is a great untruth told to a gullible lay audience. We have no clue or at least no certain knowledge about almost anything in life’s history. This does not necessarily mean that “God diddit,” but it also means that nothing has been refuted and we should not a priori exclude alternative explanations like intelligent agency. Keeping an open mind is a wise approach that is unfortunately very much ignored by modern science. True skeptics should question everything, and not just everything apart from Darwinism and materialism. I wholeheartedly endorse the credo to follow the evidence wherever it leads.
For this reason, I currently and provisionally still think that the total evidence of all lines of data favors common descent as the most parsimonious and most elegant explanation. However, I definitely remain open (and now more sympathetic) to alternatives like progressive creation combined with other explanations for the pattern of biological similarities such as Winston Ewert’s dependency graph hypothesis (Ewert 2018, 2023, Miller 2018, 2023, Reeves 2022; also see this website), which is based on an analogy to object-oriented programming, or my own suggestion of a maximization of information content as a design principle based on pattern cladistic arguments (Bechly & Meyer 2017). Incidentally, my main quibble with Ewert’s interesting approach was that it does not allow for similarly precise successful predictions as the common descent hypothesis. Looks like I have to reconsider my stance. And if even more conflicting evidence should ultimately lead me away from the paradigm of common descent, then so be it. In any case, the accumulating anomalies, incongruences, conflicting data, and other problems certainly suggest that something very different has driven the history on life on Earth than the unguided process imagined by Charles Darwin and his modern followers.
References
- Alberti G & Michalik P 2004. Feinstrukturelle Aspekte der Fortpflanzungssysteme von Spinnentieren (Arachnida). Denisia 12, 1–62. https://www.zobodat.at/pdf/DENISIA_0012_0001-0062.pdf
- Anderson EP, Schiffbauer JD, Jacquet SM, Lamsdell JC, Kluessendorf J & Mikulic DG 2021. Stranger than a scorpion: a reassessment of Parioscorpio venator, a problematic arthropod from the Llandoverian Waukesha Lagerstätte. Palaeontology 64(3), 429–474. DOI: https://doi.org/10.1111/pala.12534
- Aria C & Caron J-B 2019. A Middle Cambrian arthropod with chelicerae and proto-book gills. Nature 573, 586–589. DOI: https://doi.org/10.1038/s41586-019-1525-4
- Ballesteros JA, Santibáñez López CE, Kováč Ľ, Gavish-Regev E & Sharma PP 2019 Ordered phylogenomic subsampling enables diagnosis of systematic errors in the placement of the enigmatic arachnid order Palpigradi. Proceedings of the Royal Society B 286: 20192426, 1–9. DOI: https://doi.org/10.1098/rspb.2019.2426
- Ballesteros JA et al. 2022. Comprehensive Species Sampling and Sophisticated Algorithmic Approaches Refute the Monophyly of Arachnida. Molecular Biology and Evolution 39(2): msac021, 1–15. DOI: https://doi.org/10.1093/molbev/msac021
- Ballesteros JA & Sharma PP 2019. A critical appraisal of the placement of Xiphosura (Chelicerata) with account of known sources of phylogenetic error. Systematic Biology 68(6), 896–917. DOI: https://doi.org/10.1093/sysbio/syz011
- Ban X-C, Shao Z-K, Wu L-J, Sun J-T & Xue X-F 2022. Highly diversified mitochondrial genomes provide new evidence for interordinal relationships in the Arachnida. Cladistics 38(4), 452-464. DOI: https://doi.org/10.1111/cla.12504
- Bechly G 2020. The Oldest Scorpion and the Decadence of Evolutionary Science. Evolution News January 21, 2020. https://evolutionnews.org/2020/01/the-oldest-scorpion-and-the-decadence-of-evolutionary-science/
- Bechly G 2021. Does the Fossil Record Demonstrate Darwinian Evolution? Chapter 31, pp 345–356 in: Dembski WA, Luskin C & Holden JM (eds). The Comprehensive Guide to Science and Faith. Harvest House, Eugene (OR), 656. pp. https://www.discovery.org/b/comprehensive-guide-to-science-and-faith/
- Bechly G 2023a. The Fossil Record. Capter 7, pp. 75–90 in: Gauger A (ed.). God’s Grandeur – The Catholic Case for Intelligent Design. Sophia Institute Press, Manchester (NH), 339 pp. https://www.discovery.org/_/gods-grandeur/
- Bechly G 2023b. Fossil Friday: How the Caterpillar Got Its Legs, or Not. Evolution News November 10, 2023. https://evolutionnews.org/2023/11/fossil-friday-how-the-caterpillar-got-its-legs-or-not/
- Bechly G & Meyer SC 2017. The Fossil Record and Universal Common Ancestry. Chapter 10, pp 331–361 in: Moreland JP, Meyer SC, Shaw C, Gauger AK, Grudem W (eds). Theistic Evolution: A Scientific, Philosophical, and Theological Critique. Wheaton (IL): Crossway, 1008 pp. https://www.crossway.org/books/theistic-evolution-case/
- Benton MJ & Hitchin R 1997. Congruence between phylogenetic and stratigraphic data on the history of life. Proceedings of the Royal Society B 264(1383), 885–890. DOI: https://doi.org/10.1098/rspb.1997.0123
- Bicknell RDC, Lustri L & Birch SA 2019. Revision of “Bellinurus” carteri (Chelicerata: Xiphosura) from the Late Devonian of Pennsylvania, USA. Comptes Rendus Palevol 18(8), 967–976. DOI: https://doi.org/10.1016/j.crpv.2019.08.002
- Bond JE, Garrison NL, Hamilton CA, Godwin RL, Hedin M & Agnarsson I 2014. Phylogenomics Resolves a Spider Backbone Phylogeny and Rejects a Prevailing Paradigm for Orb Web Evolution. Current Biology 24(15), 1765–1771. DOI: https://doi.org/10.1016/j.cub.2014.06.034
- Braddy SJ, Aldridge RJ, Gabbott SE & Theron JN 1999. Lamellate book-gills in a late Ordovician eurypterid from the Soom Shale, SouthAfrica: support for a eurypterid-scorpion clade. Lethaia 32(1), 72-74. DOI: https://doi.org/10.1111/j.1502-3931.1999.tb00582.x
- Brasier M, Cotton L & Yenney I 2009. First report of amber with spider webs and microbial inclusions from the earliest Cretaceous ( c. 140 Ma) of Hastings, Sussex. Journal of the Geological Society 166, 989-997. DOI: https://doi.org/10.1144/0016-76492008-158
- Brunnetta L & Craig CL 2010. Spider Silk: Evolution and 400 Million Years of Spinning, Waiting, Snagging, and Mating. Yale University Press, New Haven (CT), 320 pp. https://books.google.de/books?id=JU0w9HVElf8C
- Cheng XD, Liu SH, Huang WJ, Liu L, Li HM & Li YX 2019. Retracted: A new species of Mongolarachnidae from the Yixian Formation of Western Liaoning China. Acta Geologica Sinica 93(1), 227–228. DOI: https://doi.org/10.1111/1755-6724.13780 (retracted article)
- Clyde WS & Fisher DC 1997. Comparing the fit of stratigraphic and morphologic data in phylogenetic analysis. Paleobiology 23(1), 1–19. DOI: https://doi.org/10.1017/S0094837300016614
- Cotton TJ / Braddy SJ 2004. The phylogeny of arachnomorph arthropods and the origin of the Chelicerata. Transactions of the Royal Society of Edinburgh: Earth Sciences 94(3), 169–193. DOI: https://doi.org/10.1017/S0263593300000596
- Damen WG, Saridaki T & Averof M 2002. Diverse adaptations of an ancestral gill: a common evolutionary origin for wings, breathing organs, and spinnerets. Current Biology 12(19), 1711–1716. DOI: https://doi.org/10.1016/S0960-9822(02)01126-0
- Delclòs X, Nel A, Azar D, Bechly G, Dunlop JA, Engel MS & Heads S 2008. The enigmatic Mesozoic insect taxon Chresmodidae (Polyneoptera): New palaeobiological and phylogenetic data, with the description of a new species from the Lower Cretaceous of Brazil. Neues Jahrbuch für Geologie und Paläontologie – Abhandlungen 247(3), 353–381. DOI: https://doi.org/10.1127/0077-7749/2008/0247-0353
- Dunlop JA 1997. Palaeozoic arachnids and their significance for arachnid phylogeny. Proceedings of the 16th European Colloquium of Arachnology, 65–82. https://www.european-arachnology.org/esa/wp-content/uploads/2015/08/065-082_Dunlop.pdf
- Dunlop JA 2010. Geological history and phylogeny of Chelicerata. Arthropod Structure & Development 39(2), 124–142. DOI: https://doi.org/10.1016/j.asd.2010.01.003
- Dunlop JA 2018. Systematics of the Coal Measures whip spiders (Arachnida: Amblypygi). Zoologischer Anzeiger 273, 14–22. DOI: https://doi.org/10.1016/j.jcz.2017.11.004
- Dunlop JA 2020. Evolution: A Breath of Fresh Air for Eurypterids. Current Biology 30(21), R1304–R1306. DOI: https://doi.org/10.1016/j.cub.2020.09.052
- Dunlop JA 2022. Spider Origins: a Palaeontological Perspective. Arachnology 19(SP1), 182–190. DOI: https://doi.org/10.13156/arac.2022.19.sp1.182
- Dunlop JA 2023. The first Palaeozoic spider (Arachnida: Araneae) from Germany. Paläontologische Zeitschrift PalZ 97, 497–504. DOI: https://doi.org/10.1007/s12542-023-00657-7
- Dunlop JA & Arango CP 2004. Pycnogonid affinities: a review. Journal of Zoological Systematics and Evolutionary Research 43(1), 8–21: DOI: https://doi.org/10.1111/j.1439-0469.2004.00284.x
- Dunlop JA & Bechly G (2015): Kieferklauenträger (Chelicerata). pp. 292–298 in: Arratia G, Schultze HP, Tischlinger H & Viohl G (eds). Solnhofen – Ein Fenster in die Jurazeit. 2 vols. Pfeil Verlag, Munich (DE), 620 pp. https://pfeil-verlag.de/publikationen/solnhofen-ein-fenster-in-die-jurazeit/
- Dunlop JA & Braddy SJ 2001. Scorpions and their sistergroup relationships. pp. 1–24 in: Fet V & Selden PA (eds). Scorpions 2001. In Memoriam Gary A. Polis. The British Arachnological Society: Burnam Beeches.
- Dunlop JA & Selden PA 1998. The early history and phylogeny of the chelicerates. Chapter 17, pp. 221–235 in: Fortey RA & Thomas RH (eds). Arthropod Relationships. Systematics Association Special Volume Series 55. Chapman & Hall, London (UK), xii+383 pp. DOI: https://doi.org/10.1007/978-94-011-4904-4_17
- Dunlop JA & Webster M 1999. Fossil Evidence, Terrestrialization and Arachnid Phylogeny. in: Proceedings of the XIV International Congress of Arachnology and a Symposium on Spiders in Agroecosystems. The Journal of Arachnology 27(1), 86–93. JSTOR: https://www.jstor.org/stable/3705969
- Dunlop JA, Tetlie OE & Prendini L 2008. Reinterpretation of the Silurian Scorpion Proscorpius osborni (Whitfield): Integrating Data from Palaeozoic and Recent Scorpions. Palaeontology 51(2), 303–320. DOI: https://doi.org/10.1111/j.1475-4983.2007.00749.x
- Dunlop JA, Scholtz G & Selden PA 2013. Water-to-Land Transitions. Chapter 16, pp. 417–439 in: Minelli A, Boxshall G & Fusco G (eds). Arthropod Biology and Evolution. Springer: Berlin / Heidelberg (DE), ix+532 pp. DOI: https://doi.org/10.1007/978-3-642-36160-9_16
- Dunlop JA, Borner J, Burmester T. 2014. Phylogeny of the Chelicerates: Morphological and Molecular Evidence. Chapter 16, pp. 399–412 in: Wägele JW & Bartolomaeus T (eds). Deep Metazoan Phylogeny: The Backbone of the Tree of lLfe. De Gruyter, Berlin (DE), xxv+736 pp. DOI: https://doi.org/10.1515/9783110277524.399
- Dunlop JA, Bird TL, Brookhart JO & Bechly, G. 2015a. A camel spider from Cretaceous Burmese amber. Cretaceous Research 56, 265–273. DOI: https://doi.org/10.1016/j.cretres.2015.05.003
- Dunlop JA, Penney D, Jekel D 2015b. A summary list of fossil spiders and their relatives. in: World Spider Catalog. Natural History Museum Bern, Bern (CH). https://wsc.nmbe.ch/resources/fossils/Fossils16.0.pdf
- Economist 2018. A strange fossil spider. Or maybe not. The Economist February 8, 2018. https://www.economist.com/science-and-technology/2018/02/08/a-strange-fossil-spider-or-maybe-not?fsrc=gp_en
- Ewert W 2018. The dependency graph of life. BIO-Complexity 2018(3), 1–27. DOI: https://doi.org/10.5048/BIO-C.2018.3
- Ewert W 2023. AminoGraph Analysis of the Auditory Protein Prestin From Bats and Whales Reveals a Dependency-Graph Signal That Is Missed by the Standard Convergence Model. BIO-Complexity 2023(1), 1–15. DOI: https://doi.org/10.5048/BIO-C.2023.1
- Funnell R 2023. 310-Million-Year-Old Fossil Spider Is The Oldest Ever Found In Germany. IFLScience July 28, 2023. https://www.iflscience.com/310-million-year-old-fossil-spider-is-the-oldest-ever-found-in-germany-70045
- Garwood RJ & Dunlop J 2014. Three-dimensional reconstruction and the phylogeny of extinct chelicerate orders. PeerJ 2: e641, 1–33. DOI: https://doi.org/10.7717/peerj.641
- Garwood RJ & Dunlop JA 2023. Consensus and conflict in studies of chelicerate fossils and phylogeny. Arachnologische Mitteilungen (accepted). https://research.manchester.ac.uk/en/publications/consensus-and-conflict-in-studies-of-chelicerate-fossils-and-phyl
- Garwood RJ, Dunlop JA, Selden PA, Spencer ART, Atwood RC, Vo NT & Drakopoulos Michael 2016. Almost a spider: a 305-million-year-old fossil arachnid and spider origins. Proceedings of the Royal Society B 283(1827): 20160125, 1–8. DOI: https://doi.org/10.1098/rspb.2016.0125
- Garwood RJ, Dunlop JA, Knecht BJ & Hegna TA 2017. The phylogeny of fossil whip spiders. BMC Evolutionary Biology 17: 105, 1–14. DOI: https://doi.org/10.1186/s12862-017-0931-1
- Giribet G 2018. Current Views on Chelicerate phylogeny— A Tribute to Peter Weygoldt. Zoologischer Anzeiger 273, 7–13. DOI: https://doi.org/10.1016/j.jcz.2018.01.004
- Giribet G & Edgecombe GD 2019. The Phylogeny and Evolutionary History of Arthropods. Current Biology 29(12), R592–R602. DOI: https://doi.org/10.1016/j.cub.2019.04.057
- Giribet G, Edgecombe GD, Wheeler WC & Babbitt C 2002. Phylogeny and Systematic Position of Opiliones: A Combined Analysis of Chelicerate Relationships Using Morphological and Molecular Data. Cladistics 18(1), 5–70. DOI: https://doi.org/10.1111/j.1096-0031.2002.tb00140.x
- Göpel T & Wirkner CS 2015. An “ancient” complexity? Evolutionary morphology of the circulatory system in Xiphosura. Zoology 118(4), 221–238. DOI: https://doi.org/10.1016/j.zool.2014.12.004
- Grasshoff M 1978. A model of the evolution of the main chelicerate groups. Symposium of the Zoological Society of London 42, 273–284.
- Gray M 2018. Spider origins. Australian Museum website November 26, 2018. https://australian.museum/learn/animals/spiders/spider-origins/
- Harvey MS 2002. The neglected cousins: what do we know about the smaller arachnid orders? The Journal of Arachnology 30(2), 357–372. DOI: https://doi.org/10.1636/0161-8202(2002)030[0357:TNCWDW]2.0.CO;2
- Hilbrant M 2008. Development and Evolution of the Spider Silk Production System. Ph.D. thesis, University of Cologne: Cologne (DE), vi+ 144 pp. https://kups.ub.uni-koeln.de/2795/
- Howard RJ, Edgecombe GD, Legg DA, Pisani D & Lozano-Fernandez J 2019. Exploring the evolution and terrestrialization of scorpions (Arachnida: Scorpiones) with rocks and clocks. Organisms Diversity & Evolution 19(1), 71–86. DOI: https://doi.org/10.1007/s13127-019-00390-7
- Howard RJ, Puttick MN, Edgecombe GD & Lozano-Fernandez J 2020. Arachnid monophyly: Morphological, palaeontological and molecular support for a single terrestrialization within Chelicerata. Arthropod Structure & Development 59: 100997, 1-14. DOI: https://doi.org/10.1016/j.asd.2020.100997
- Huang D, Hormiga G, Cai C, Su Y, Yin Z, Xia F & Giribet G 2018. Origin of Spiders and Their Spinning Organs Illuminated by Mid-Cretaceous Amber Fossils. Nature Ecology & Evolution 2(4), 623–627. DOI: https://doi.org/10.1038/s41559-018-0475-9
- Hünicken MA 1980. A giant fossil spider (Megarachne servinei) from Bajo de Véliz, Upper Carboniferous, Argentina. Boletin de la Academia Nacional de Ciencias (Cordoba) 53(3-4), 317–341.
- Kamenz C 2009. Book-lung morphology: implications for arachnid phylogeny (Arachnida, Chelicerata). PhD thesis, Humboldt University, Berlin (DE), 234 pp. DOI: https://doi.org/10.18452/16070
- Kamenz C, Dunlop JA, Scholtz G, Kerp H & Hass H 2008. Microanatomy of Early Devonian book lungs. Biology Letters 4(2), 212–215. DOI: https://doi.org/10.1098/rsbl.2007.0597
- Kjellesvig-Waering EN 1986: A Restudy of the Fossil Scorpionida of the World. Palaeontographica Americana 55, 1–287.
- Kraus O 1976. Zur phylogenetischen Stellung und Evolution der Chelicerata. Entomologica Germanica 3(1-2), 1–12. DOI: https://doi.org/10.1127/entom.germ/3/1976/1
- Kühl G, Bergmann A, Dunlop J, Garwood RJ & Rust J 2012. Redescription and Palaeobiology of Palaeoscorpius devonicus Lehmann, 1944 from the Lower Devonian Hunsrück Slate of Germany. Palaeontology 55(4), 775–787. DOI: https://doi.org/10.1111/j.1475-4983.2012.01152.x
- Kuntner M 2022. The seven grand challenges in arachnid science. Frontiers in Arachnid Science 1, 1–12. DOI: https://doi.org/10.3389/frchs.2022.1082700
- Lamsdell JC 2012. Revised systematics of Palaeozoic ‘horseshoe crabs’ and the myth of monophyletic Xiphosura. Zoological Journal of the Linnean Society 167(1), 1–27. DOI: https://doi.org/10.1111/j.1096-3642.2012.00874.x
- Lamsdell JC 2016. Horseshoe crab phylogeny and independent colonizations of fresh water: Ecological invasion as a driver for morphological innovation. Palaeontology 59(2), 181–194. DOI: https://doi.org/10.1111/pala.12220
- Lamsdell JC, Briggs DEG, Liu HP, Witzke BJ & McKay RM 2015. A new Ordovician arthropod from the Winneshiek Lagerstätte of Iowa (USA) reveals the ground plan of eurypterids and chasmataspidids. Naturwissenschaften / The Science of Nature 102(9-10), 63. DOI: https://doi.org/10.1007/s00114-015-1312-5
- Lamsdell JC, McCoy VE, Perron-Feller OA & Hopkins MJ 2020. Air Breathing in an Exceptionally Preserved 340-Million-Year-Old Sea Scorpion. Current Biology 30(21), 4316-4321. DOI: https://doi.org/10.1016/j.cub.2020.08.034
- Lauterbach K-E 1980. Schlüsselereignisse in der Evolution des Grundplans der Arachnata (Arthropoda). Abhandlungen des Naturwissenschaftlichen Vereins in Hamburg (NF) 23, 163–327. [In German]
- Legg DA, Sutton MD & Edgecombe DA 2013. Arthropod fossil data increase congruence of morphological and molecular phylogenies. Nature Communications 4: 2485, 1-7. DOI: https://doi.org/10.1038/ncomms3485
- Lozano-Fernandez J, Tanner A, Giacomelli M, Carton R, Vinther J, Edgecombe GD & Pisani D 2019. Increasing species sampling in chelicerate genomic-scale datasets provides support for monophyly of Acari and Arachnida. Nature Communications 10(1); 2295, 1–8. DOI: https://doi.org/10.1038/s41467-019-10244-7
- Lozano-Fernandez J, Tanner AR, Puttick MN, Vinther J, Edgecombe GD & Pisani D 2020. A Cambrian–Ordovician Terrestrialization of Arachnids. Frontiers in Genetics 11: 182, 1–11. DOI: https://doi.org/10.3389/fgene.2020.00182
- Luskin C 2023. Intelligent Design Passes the Dawkins Test. Evolution News May 15, 2023. https://evolutionnews.org/2023/05/intelligent-design-passes-the-dawkins-test/
- Mariano-Martins P, Lo-Man-Hung N & Teixeira Torres T 2020. Evolution of Spiders and Silk Spinning: Mini Review of the Morphology, Evolution, and Development of Spiders’ Spinnerets. Frontiers in Ecology and Evolution 8: 109, 1–6. DOI: https://doi.org/10.3389/fevo.2020.00109
- McLatchie J 2012. Jerry Coyne’s Chapter on the Fossil Record Fails to Show “Why Evolution Is True”. Evolution News December 4, 2012 https://evolutionnews.org/2012/12/jerry_coynes_c/
- Miether ST & Dunlop JA 2016. Lateral eye evolution in the arachnids. Arachnology 17(2), 103–119. DOI: https://doi.org/10.13156/arac.2006.17.2.103
- Miller B 2018. BIO-Complexity Presents Better Model than Common Ancestry for Explaining Pattern of Nature. Evolution News July 19, 2018. https://evolutionnews.org/2018/07/bio-complexity-presents-a-better-model-than-common-ancestry-for-explaining-the-pattern-of-nature/
- Miller B 2023. From Winston Ewert, New Peer-Reviewed Paper on Dependency Graph Model. Evolution News August 23, 2023. https://evolutionnews.org/2023/08/new-peer-reviewed-paper-on-dependency-graph-model/
- Neethling JA 2021. Ancient and Extant – The History of Arachnids. National Museum Publications July 19, 2001. https://nationalmuseumpublications.co.za/ancient-and-extant-the-history-of-arachnids/
- Noah KE, Hao J, Li L, Sun X, Foley B, Yang Q & Xia X 2020. Major Revisions in Arthropod Phylogeny Through Improved Supermatrix, With Support for Two Possible Waves of Land Invasion by Chelicerates. Evolutionary Bioinformatics 16, 1–12. DOI: https://doi.org/10.1177/1176934320903735
- Norell MA & Novacek MJ 1992. Congruence between superpositional and phylogenetic patterns: Comparing cladistic patterns with fossil records. Cladistics 8(4), 319–337. DOI: https://doi.org/10.1111/j.1096-0031.1992.tb00074.x
- Ontano AZ, Gainett G, Aharon S, Ballesteros JA, Benavides LR et al. 2021. Taxonomic Sampling and Rare Genomic Changes Overcome Long-Branch Attraction in the Phylogenetic Placement of Pseudoscorpions. Molecular Biology and Evolution 38(6), 2446–2467. DOI: https://doi.org/10.1093/molbev/msab038
- Ontano AZ, Steiner HG & Sharma PP. 2022. How many long branch orders occur in Chelicerata? Opposing effects of Palpigradi and Opilioacariformes on phylogenetic stability. Molecular Phylogenetics and Evolution 168: 107378, 1–9. DOI: https://doi.org/10.1016/j.ympev.2021.107378
- Pappas S 2016. 305-Million-Year-Old ‘Almost Spider’ Unlocks Arachnid History. Scientific American March 31, 2016. https://www.scientificamerican.com/article/305-million-year-old-almost-spider-unlocks-arachnid-history/
- Paulus HF 2004. Einiges zur Stammesgeschichte der Spinnentiere (Arthropoda, Chelicerata). Denisia (NS)14, 547–574. [In German] https://www.zobodat.at/pdf/DENISIA_0012_0547-0574.pdf
- Pechmann M & Prpic NM 2009. Appendage patterning in the South American bird spider Acanthoscurria geniculata (Araneae: Mygalomorphae). Development Genes and Evolution 219, 189–198. DOI: https://doi.org/10.1007/s00427-009-0279-7
- Peñalver E, Grimaldi DA & Delclòs X 2006. Early Cretaceous Spider Web with Its Prey. Science 312(5781), 1761. DOI: https://doi.org/10.1126/science.1126628
- Penney D & Ortuño VM 2006. Oldest true orb-weaving spider (Araneae: Araneidae). Biology Letters 2(3), 1–4. DOI: https://doi.org/10.1098/rsbl.2006.0506
- Pepato AR, da Rocha CE & Dunlop JA 2010. Phylogenetic position of the acariform mites: sensitivity to homology assessment under total evidence. BMC Evolutionary Biology 10(1): 235, 1–23. DOI: https://doi.org/10.1186/1471-2148-10-235
- Pocock RI 1911. A Monograph of the Terrestrial Carboniferous Arachnida of Great Britain. Monographs of the Palaeontographical Society 64(315), 1–84. DOI: https://doi.org/10.1080/02693445.1911.12035553
- Pollitt JR, Braddy SJ & Dunlop JA 2004. The phylogenetic position of the extinct arachnid order Phalangiotarbida Haase, 1890, with reference to the fauna from the Writhlington Geological Nature Reserve (Somerset, UK). Transactions of the Royal Society of Edinburgh: Earth Sciences 94(3), 243–259. DOI: https://doi.org/10.1017/S0263593300000651
- Poschmann M & Dunlop JA 2006. A new sea spider (Arthropoda: Pycnogonida) with a flagelliform telson from the Lower Devonian Hunsrück Slate, Germany. Palaeontology 49(5), 983–989. DOI: https://doi.org/10.1111/j.1475-4983.2006.00583.x
- Reeves E 2022. Yes, Winston Ewert’s Dependency Graph Is a Real Model. Evolution News August 9, 2022. https://evolutionnews.org/2022/08/yes-winston-ewerts-dependency-graph-is-a-real-model/
- Regier JC & Schultz JW 1998. Molecular Phylogeny of Arthropods and the Significance of the Cambrian “Explosion” for Molecular Systematics. American Zoologist 38(6), 918–928. DOI: https://doi.org/10.1093/icb/38.6.918
- Regier JC, Shultz JW, Zwick A, Hussey A, Ball B, Wetzer R, Martin JW & Cunningham CW 2010. Arthropod relationships revealed by phylogenomic analysis of nuclear protein-coding sequences. Nature 463(7284), 1079–1083. DOI: https://doi.org/10.1038/nature08742
- Saint Martin J-P, Saint Martin S, Bolte S & Néraudeau D 2014. Spider web in Late Cretaceous French amber (Vendée): The contribution of 3D image microscopy. Comptes Rendus Palevol 13(5), 463–472. DOI: https://doi.org/10.1016/j.crpv.2014.03.005
- Schoenemann B, Poschmann M & Clarkson ENK 2019. Insights into the 400 million-year-old eyes of giant sea scorpions (Eurypterida) suggest the structure of Palaeozoic compound eyes. Scientific Reports 9(1): 17797, 1–10. DOI: https://doi.org/10.1038/s41598-019-53590-8
- Scholtz G & Kamenz C 2006. The book lungs of Scorpiones and Tetrapulmonata (Chelicerata, Arachnida): Evidence for homology and a single terrestrialisation event of a common arachnid ancestor. Zoology 109(1), 2–13. DOI: https://doi.org/10.1016/j.zool.2005.06.003
- Schwager EE, Schönauer A, Leite DJ, Sharma PP, McGregor AP 2015. Chelicerata. pp. 99–139 in: Wanninger A (ed.). Evolutionary Developmental Biology of Invertebrates 3: Ecdysozoa I: Non-Tetraconata. Springer: Vienna (AT), vii+200 pp. DOI: https://doi.org/10.1007/978-3-7091-1865-8_5
- Selden PA 1990. Fossil history of the arachnids. Newsletter of the British Arachnological Society 58, 4–6. http://www.paulselden.net/uploads/7/5/3/2/7532217/newsletterfossilarachnids.pdf
- Selden PA 1996. First fossil mesothele spider from the Carboniferous of France. Revue Suisse de Zoologie hors série, 585–596. http://www.paulselden.net/uploads/7/5/3/2/7532217/eothele.pdf
- Selden PA 2021. New spiders (Araneae: Mesothelae), from the Carboniferous of New Mexico and England, and a review of Paleozoic Araneae. In: Lucas SG, DiMichele WA & Allen BD (eds). Kinney Brick Quarry Lagerstätte. New Mexico Museum of Natural History and Science Bulletin 84, 317–358. https://www.researchgate.net/publication/348836567
- Selden PA & Dunlop JA 1998. Fossil Taxa and Relationships of Chelicerates. Chapter 7, pp. 303–331 in: Edgecombe GD (ed.). Arthropod Fossils and Phylogeny. Columbia University Press: New York (NY), 347 pp.
- Selden PA & Jeram AJ 1989. Palaeophysiology of terrestrialisation in the Chelicerata. Transactions of The Royal Society of Edinburgh: Earth Sciences 80(3-4), 303–310. DOI: https://doi.org/10.1017/S0263593300028741
- Selden PA & Penney D 2010. Fossil spiders. Biological Reviews 85(1), 171–206. DOI: https://doi.org/10.1111/j.1469-185X.2009.00099.x
- Selden PA, Shear WA & Bonamo PM 1991. A spider and other arachnids from the Devonian of New York, and reinterpretations of Devonian Araneae. Palaeontology 34(2), 241–281. https://www.palass.org/publications/palaeontology-journal/archive/34/2/article_pp241-281
- Selden PA, Corronca JA & Hünicken MA 2005. The true identity of the supposed giant fossil spider Megarachne. Biology Letters 1(1), 44–48. DOI: https://doi.org/10.1098/rsbl.2004.0272
- Selden PA, Shear WA & Sutton MD 2008. Fossil evidence for the origin of spider spinnerets, and a proposed arachnid order. PNAS 105(52), 20781–20785. DOI: https://doi.org/10.1073/pnas.0809174106
- Selden PA, Shcherbakov DE, Dunlop JA, Eskov KY 2014. Arachnids from the Carboniferous of Russia and Ukraine, and the Permian of Kazakhstan. Paläontologische Zeitschrift 88(3), 297–307. DOI: https://doi.org/10.1007/s12542-013-0198-9
- Selden PA, Lamsdell JC & Qi L 2015. An unusual euchelicerate linking horseshoe crabs and eurypterids, from the Lower Devonian (Lochkovian) of Yunnan, China. Zoologica Scripta 44(6), 645–652. DOI: https://doi.org/10.1111/zsc.12124
- Selden PA, Dunlop JA, Garwood RJ 2016. Carboniferous araneomorph spiders reinterpreted as long-bodied harvestmen. Journal of Systematic Palaeontology 14(2), 127–137. DOI: https://doi.org/10.1080/14772019.2015.1018969
- Selden PA, Olcott AN, Downen MR, Ren D, Shih C & Cheng X 2019. The supposed giant spider Mongolarachne chaoyangensis, from the Cretaceous Yixian Formation of China, is a crayfish. Palaeoentomology 002(5), 515–522. DOI: https://doi.org/10.11646/palaeoentomology.2.5.15
- Sharma PP, Kaluziak ST, Pérez-Porro AR, González VL, Hormiga G, Wheeler WC & Giribet G. 2014. Phylogenomic Interrogation of Arachnida Reveals Systemic Conflicts in Phylogenetic Signal. Molecular Biology and Evolution 31(11), 2963–2984. DOI: https://doi.org/10.1093/molbev/msu235
- Sharma PP, Ballesteros JA & Santibáñez-López CE 2021. What Is an “Arachnid”? Consensus, Consilience, and Confirmation Bias in the Phylogenetics of Chelicerata. Diversity 13(11): 568, 1–33. DOI: https://doi.org/10.3390/d13110568
- Scholtz G & Kamenz C 2006. The Book Lungs of Scorpiones and Tetrapulmonata (Chelicerata, Arachnida): Evidence for Homology and a Single Terrestrialisation Event of a Common Arachnid Ancestor. Zoology 109(1), 2–13. DOI: https://doi.org/10.1016/j.zool.2005.06.003
- Shear WA, Palmer JM, Coddington JA & Bonamo PM 1989. A Devonian Spinneret: Early Evidence of Spiders and Silk Use. Science 246(4929), 479–481. DOI: https://doi.org/10.1126/science.246.4929.479
- Shingate P, Ravi V, Prasad A, Tay B-H, Garg KM, Chattopadhyay B, Yap L-M, Rheindt FE & Venkatesh B 2020. Chromosome- level assembly of the horseshoe crab genome provides insights into its genome evolution. Nature Communications 11: 2322, 1–13. DOI: https://doi.org/10.1038/s41467-020-16180-1
- Shoemaker AK, Setton EVW & Sharma PP 2017. Differential expression of dachshund in epipod and telopod derivatives suggests non-homology of crustacean gills and spider spinnerets. 2017 Annual Meeting of the Society for Integrative and Comparative Biology January 6, 2017. https://sicb.org/abstracts/differential-expression-of-dachshund-in-epipod-and-telopod-derivatives-suggests-non-homology-of-crustacean-gills-and-spider-spinnerets/
- Shultz JW 1989. Morphology of locomotor appendages in Arachnida: evolutionary trends and phylogenetic implications. Zoological Journal of the Linnean Society 97(1), 1–56. DOI: https://doi.org/https://doi.org/10.1111/j.1096-3642.1989.tb00552.x
- Shultz JW 1990. Evolutionary Morphology and Phylogeny of Arachnida. Cladistics 6(1), 1–38. DOI: https://doi.org/10.1111/j.1096-0031.1990.tb00523.x
- Shultz JW 1994. The limits of stratigraphic evidence in assessing phylogenetic hypotheses of recent arachnids. Journal of Arachnology 22(2), 169–172. JSTOR: https://www.jstor.org/stable/3705614
- Shultz JW 1999. Muscular anatomy of a whipspider, Phrynus longipes (Pocock) (Arachnida: Amblypygi), and its evolutionary significance. Zoological Journal of the Linnean Society 126(1), 81–116. DOI: https://doi.org/10.1111/j.1096-3642.1999.tb00608.x
- Shultz JW 2007. A phylogenetic analysis of the arachnid orders based on morphological characters. Zoological Journal of the Linnean Society 150(2), 221–265. DOI: https://doi.org/10.1111/j.1096-3642.2007.00284.x
- Smith EL 1990. An arthropod morphologist looks at six hundred million years of chelicerate evolution. Newsletter of the American Arachnological Society 42, 3–5. https://www.americanarachnology.org/fileadmin/documents/am_arachnol_newsletter/AmerArachnol42_Oct_1990.pdf
- Snodgrass RE 1938. Evolution of the Annelida, Onychophora and Arthropoda. Smithsonian Miscellaneous Collections 97(6), 1–159. https://repository.si.edu/handle/10088/23666
- Starobogatov YI 1990. Sistema i filogeniya nizshikh khelit-serovykh (analiz morfologii paleozoysikh grupp). [The systematics and phylogeny of the lower chelicerates (a morphological analysis of the Paleozoic groups)]. Paleontologicheskii Zhurnal 1, 4–17. [In Russian with English translation]
- Starr M 2019. A Fossil Spider Discovery Just Turned Out to Be a Crayfish With Some Legs Painted on. ScienceAlert December 20, 2019. https://www.sciencealert.com/a-fossil-has-been-stripped-of-its-classification-because-its-legs-were-literally-painted-on
- Starrett J, Derkarabetian S, Hedin M, Bryson RW Jr, McCormack JE & Faircloth BC 2016. High Phylogenetic Utility of an Ultraconserved Element Probe Set Designed for Arachnida. Molecular Ecology Resources 17(4), 812–823. DOI: https://doi.org/10.1111/1755-0998.12621
- Switek B 2010. Megarachne, the Giant Spider That Wasn’t. ScienceBlogs March 24, 2010. https://scienceblogs.com/laelaps/2010/03/24/megarachne-the-giant-spider-th
- University of Kansas 2018. Remarkable spider with a tail found preserved in amber after 100 million years. EurekAlert! February 5, 2018. https://www.eurekalert.org/news-releases/921832
- University of Kansas 2019. A ‘Jackalope’ of an Ancient Spider Fossil Deemed a Hoax, Unmasked as a Crayfish. The University of Kansas December 19, 2019. https://today.ku.edu/2019/12/17/‘jackalope’-ancient-spider-fossils-deemed-hoax-unmasked-crayfish
- van der Hammen L 1986. Acarological and arachnological notes. Zool. Meded. 60(14), 217–230. https://repository.naturalis.nl/pub/319156
- van der Hammen L 1989. An Introduction to Comparative Arachnology. SPB Academic Publishing: The Hague (NL), 576 pp.
- Vollrath F & Selden P 2007. The Role of Behavior in the Evolution of Spiders, Silks, and Webs. Annual Review of Ecology, Evolution, and Systematics 38, 819–846. DOI: https://doi.org/10.1146/annurev.ecolsys.37.091305.110221
- Waddington J, Rudkin DM & Dunlop JA 2015. A new mid- Silurian aquatic scorpion – one step closer to land? Biology Letters 11: 20140815, 1–4. DOI: https://doi.org/10.1098/rsbl.2014.0815
- Wang B 2018. When spiders had tails. Ecology & Evolution Community February 8, 2018. https://ecoevocommunity.nature.com/posts/30041-when-spiders-had-tails
- Wang B, Dunlop JA, Selden PA, Garwood RJ, Shear WA, Müller P & Lei X 2018. Cretaceous arachnid Chimerarachne yingi gen. et sp. nov. illuminates spider origins. Nature Ecology & Evolution 2(4), 614–622. DOI: https://doi.org/10.1038/s41559-017-0449-3
- Wendruff AJ, Babcock LE, Wirkner CS, Kluessendorf J & Mikulic DG 2020. A Silurian ancestral scorpion with fossilised internal anatomy illustrating a pathway to arachnid terrestrialisation. Scientific Reports 10: 14, 1–5. DOI: https://doi.org/10.1038/s41598-019-56010-z
- Weygoldt P 1998. Evolution and systematics of the Chelicerata. Experimental & Applied Acarology 22(2), 63–79. DOI: https://doi.org/10.1023/A:1006037525704
- Weygoldt P 1999. Evolution and systematics of the Chelicerata. pp. 1–14 in: Bruin J, van der Geest LPS & Sabelis MW (eds). Ecology and Evolution of the Acari. Series Entomologica 55. Springer, Dordrecht (NL), xii+677 pp. DOI: https://doi.org/10.1007/978-94-017-1343-6_1
- Weygoldt P & Paulus HF 1979. Untersuchungen zur Morphologie, Taxonomie und Phylogenie der Chelicerata II. Cladogramme und die Entfaltung der Chelicerata. Journal of Zoological Systematics and Evolutionary Research 17(3), 177–200. DOI: https://doi.org/10.1111/j.1439-0469.1979.tb00699.x
- Wheeler WC & Hayashi CY 1998. The phylogeny of the extant chelicerate orders. Cladistics 14(2), 173–192. DOI: https://doi.org/10.1111/j.1096-0031.1998.tb00331.x
- Wheeler WC et al. 2016. The spider tree of life: phylogeny of Araneae based on target-gene analyses from an extensive taxon sampling. Cladistics 33(6), 574–616. DOI: https://doi.org/10.1111/cla.12182
- Wolfe JM 2017. Metamorphosis Is Ancestral for Crown Euarthropods, and Evolved in the Cambrian or Earlier. Integrative & Comparative Biology 57(3), 499–509. DOI: https://doi.org/10.1093/icb/icx039
- Wunderlich J 2015. On the evolution and the classification of spiders, the Mesozoic spider faunas, and descriptions of new Cretaceous taxa mainly in amber from Myanmar (Burma) (Arachnida: Araneae). pp. 21–408 in: Wunderlich J (ed.). Beiträge zur Araneologie, Vol. 9, Mesozoic Spiders. Publishing House Joerg Wunderlich: Hirschberg (DE), 512 pp.
- Wunderlich J 2019. What is a spider? pp. 1–32 in: Wunderlich J (ed.). Beiträge zur Araneologie, Vol. 12. Publishing House Joerg Wunderlich: Hirschberg (DE). http://www.joergwunderlich.de/Downloads/Beitr._Araneol._Band_12_(2019).pdf
- Wunderlich J & Müller P 2022. Some spiders in Cretaceous amber from Myanmar (Araneida: Chimerarachnida and Araneae). pp. 119–173 in: Wunderlich J (ed.). Beiträge zur Araneologie, Vol. 15. Publishing House Joerg Wunderlich: Hirschberg (DE), 210 pp. http://www.joergwunderlich.de/Downloads/Beitr._Araneol._Band_15_(2022).pdf
- Zschokke S 2003. Spiderweb silk from the Early Cretaceous. Nature 424(6949), 636–637. DOI: https://doi.org/10.1038/424636a
- Zschokke S 2004. Glue droplets in fossil spider webs. in: Logunov DV & Penney D (eds). European Arachnology 2003. Arthropoda Selecta SI1, 367–374. https://www.researchgate.net/publication/285766549