 Evolution
Evolution
 Intelligent Design
Intelligent Design
Getting It Together: Tethers, Handshakes, and Multitaskers in the Cell
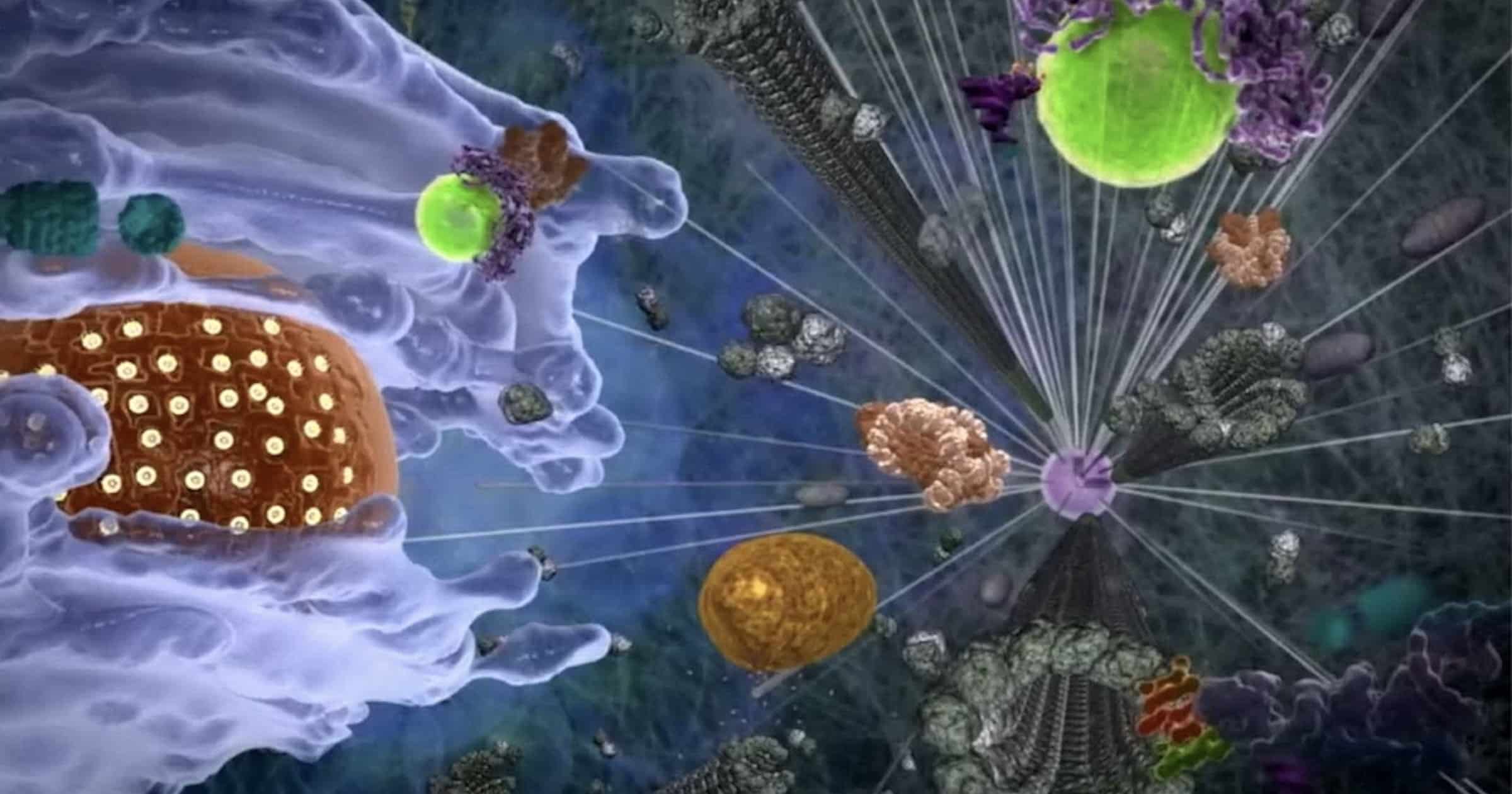
Running a cell requires coordination. How do molecules moving in the dark interior of a cell know how and when to connect? Protein tethers offer new clues, according to research at Philipps University in Marburg, Germany.
The ways that organelles and proteins connect at the right place and time are coming to light. One method is to encapsulate interacting molecules within compartments called condensates, droplets, and speckles. Like offices or cubicles where employees can talk without excess noise, these temporary spaces allow molecules to interact in peace (see “Caltech Finds Amazing Role for Noncoding DNA”).
Another method for coordination of moving parts involves tethers. Certain molecular machines use “two hands” to bring other molecules or organelles together. Visualize a person taking a stranger’s hand and using her other hand to grasp a doorknob, leading the stranger to the place he needs to be. Many protein machines have a critical binding site for their targets, but these “dual affinity” tethering machines contain two different recognition sites on different domains that recognize separate targets needing to come together. Such multitasking machines are marvelously designed to promote fellowship for effective interactions in the cellular city.
A similar phenomenon has long been known in DNA translation. A set of molecules called aminoacyl-tRNA synthetases brings dissimilar molecules together. One synthetase feels the anticodon on its matching transfer RNA (tRNA) and then puts the corresponding amino acid on the opposite end. Like a language translator, each synthetase needs to know two languages — the DNA code and the protein code — to equip the tRNA with the correct amino acid. As the activated tRNA enters the ribosome, its anticodon base pairs with the complementary codon on the messenger RNA at one end, and its amino acid fits onto the growing polypeptide chain on the other end. This is a spectacular example of double duty, multitasking know-how. But is it the only one?
Another Example of Double Duty
A team of 15 researchers publishing in PLOS Biology under lead author Elena Bittner, also from Philipps University, and colleagues at Berkeley and Howard Hughes, has just reported a case of a multitasking machine that bridges dissimilar targets — in this case, peroxisomes with mitochondria or the endoplasmic reticulum (ER). It may not be the only case of “Proteins that carry dual targeting signals [that] can act as tethers between” organelles, they say:
Peroxisomes are organelles with crucial functions in oxidative metabolism. To correctly target to peroxisomes, proteins require specialized targeting signals. A mystery in the field is the sorting of proteins that carry a targeting signal for peroxisomes and as well as for other organelles, such as mitochondria or the endoplasmic reticulum (ER). Exploring several of these proteins in fungal model systems, we observed that they can act as tethers bridging organelles together to create contact sites.
Take note that they found this in yeast, the simplest of eukaryotes.
We show that in Saccharomyces cerevisiae this mode of tethering involves the peroxisome import machinery, the ER–mitochondria encounter structure (ERMES) at mitochondria and the guided entry of tail-anchored proteins (GET) pathway at the ER.
Why is this significant?
Our findings introduce a previously unexplored concept of how dual affinity proteins can regulate organelle attachment and communication.
Previously unexplored: this sounds like a game changer. How does this “tethering” system work? After all the biochemistry work by the team is shown, demonstrating the dual-targeting capability, they illustrated it with a simplified diagram in Figure 10 in their open-access paper. As usual, even in simplified form, the system involves numerous other factors. The upshot is described as follows:
We have found that distinct proteins with targeting signals for 2 organelles can affect proximity of these organelles. This conclusion is supported by the notion that different types of dual affinity proteins can act as contact-inducing proteins (Fig 10) … Although dual affinity proteins are a challenge for maintaining organelle identity, they are ideally suited to support organelle interactions by binding to targeting factors and membrane-bound translocation machinery of different organelles. Dually targeted proteins appear to concentrate in regions of organelle contact, which may coincide with regions of reduced identity.
Within the mitochondria, we already met TIM and TOM, the channel guards who check the credentials of proteins entering the organelle’s outer and inner membranes. (The authors note that these translocase proteins are “evolutionarily conserved.”) But outside the mitochondrion, proteins needing to enter or exit have to find their way to the guards. That’s where the “dual affinity proteins” operate.
What Do the Tethers Look Like?
Ptc5 is one of these tethering proteins, one of many that “contain targeting signals for mitochondria and peroxisomes at opposite termini.” Its Peroxisome Targeting Signal (PTS) recognizes the peroxisome at one end, and its Mitochondrial Targeting Signal (MTS) recognizes TOM at the mitochondrial channel. Experimenting with mutant strains of this and associated proteins and chaperones, the researchers confirmed that Ptc5 does tether peroxisomes to mitochondria. Moreover, its activity is dependent on need. “In aggregate,” they write, these data show that tethering via dual affinity proteins is a regulated process and depends on the metabolic state of the cell.” This implies the additional capability of sensing the fluctuating metabolic need.
The authors didn’t have much to say about evolution. As usual, it involved copious amounts of speculation.
While many peroxisomal membrane proteins can target peroxisomes without transitioning through the ER, several peroxisomal membrane proteins have evolved to be synthesized in vicinity to the ER and may translocate from it.
Other than TOM and TIM being “evolutionarily conserved,” that was all they had to offer Darwin.
A New Class of Activity Coordinators
What Bittner et al. have identified is probably the trigger for a paradigm shift concerning methods that cells use to get components together.
We conclude that dually targeted cargo includes a diverse and unexpected group of tethers, which are likely to maintain contact as long as they remain accessible for targeting factors at partner organelles. Coupling of protein and membrane trafficking is a common principle in the secretory pathway and it might also occur for peroxisomes at different contact sites.
And so, what lies ahead? Design proponents in biochemistry and molecular biology, play tetherball! Here is a potentially fruitful area for new discoveries.
How dually targeted proteins and their rerouting affect the flux of molecules other than proteins, e.g., membrane lipids remains a topic for future research.

