 Evolution
Evolution
 Intelligent Design
Intelligent Design
More Scientific Problems with Paul Rimmer’s Views on Origin of Life

In an article here yesterday, I analyzed astrochemist Paul Rimmer’s commentary on the debate between James Tour and Dave Farina. I focused on the specific research Rimmer cited and how his position as an origin of life (OOL) researcher likely shapes his evaluation of its relevance to explaining how a cell could have emerged through natural processes. Here, I will describe additional serious scientific problems with Rimmer’s optimistic assessment of the state of the field.
Proposed Path to Life
Rimmer presents the stages toward life’s genesis proposed by leading origins researcher John Sutherland (Figure 1). The postulated path to life entails a continuous series of stable systems gradually increasing in complexity until an autonomous cell emerges capable of Darwinian evolution. Since Sutherland assumes that life must have originated from undirected physical processes, he believes such stable intermediates must have existed.
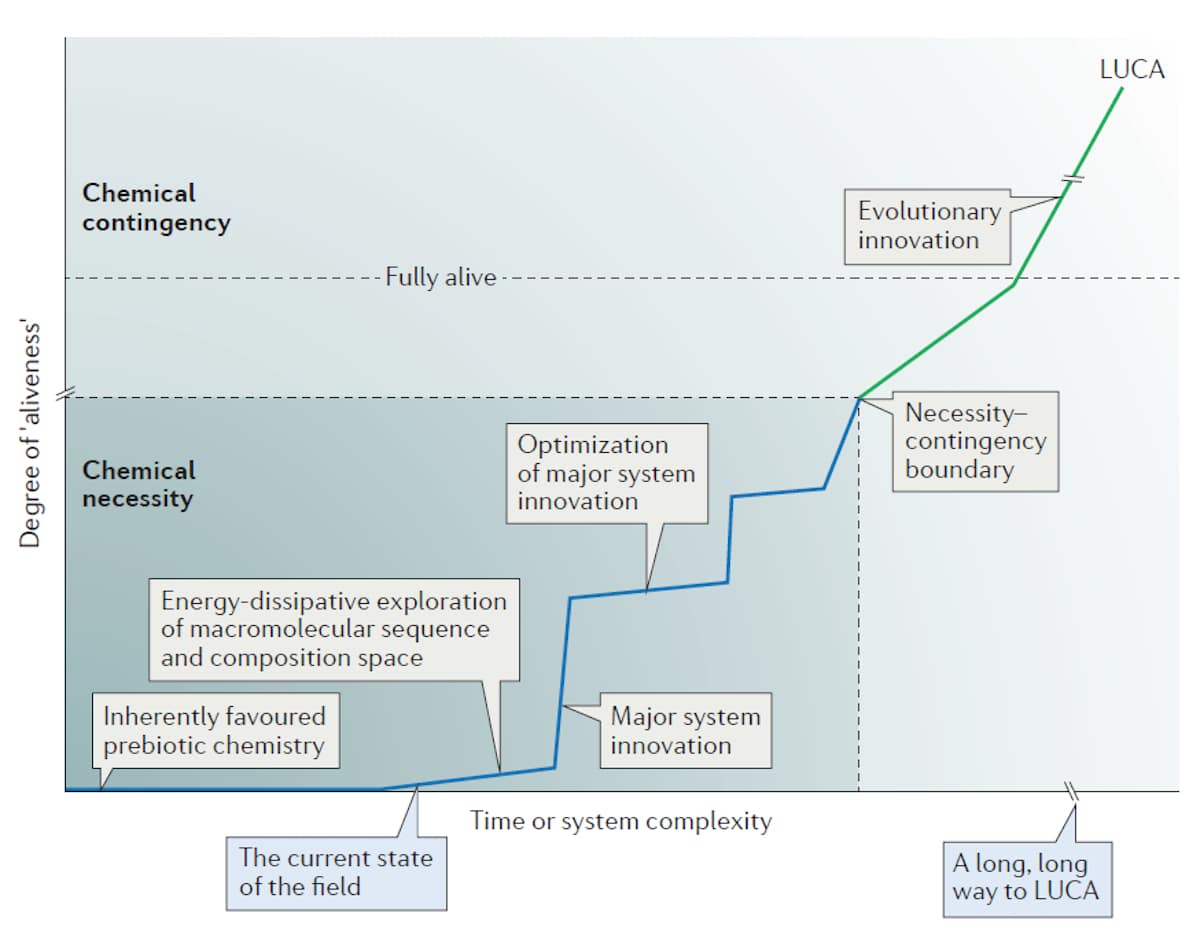
Many have attempted to justify this assumption by appealing to such researchers as Jeremy England who have demonstrated that far-from-equilibrium systems can generate self-organizational behavior such as a hurricane’s funnel cloud. Theorists propose that analogous self-organizational processes could have driven a chemical system from one stable state to the next.
The Underlying Challenge
The problem with such claims is that naturally occurring self-organization is fundamentally different from the order in living systems, and all experiments that demonstrate self-organizational behavior that is even remotely life-like must apply artificial energy sources to systems designed to properly interact with the applied energy. For instance, Bachelard et al. (2017) applied acoustic waves of specific frequencies to meniscus particles in a waveguide to generate desired patterns in the acoustic bandgap (i.e., graph of transmitted frequencies). And Sacanna et al. (2014) applied light within carefully chosen frequency ranges to asymmetric semiconductor spheres in a solution of hydrogen peroxide fuel to generate self-propulsion and crystal formation. I describe other studies in my response to Jeremy England that employed comparable investigator ingenuity. In every example, the self-organizational behavior depends on the expertise of the researchers acting to achieve desired results.
In contrast, energy sources available on the early Earth properly interact with few, if any, biologically relevant chemical reactions or physical processes. Instead, raw energy tends to break apart complex molecules (e.g., proteins and RNA) and cellular structures. Any interactions that would have allowed a chemical system to move even a tiny step toward life would have been exceedingly rare.
The same holds true for even the earliest stages of origin scenarios. Tour has detailed how research that purportedly moves a chemical system toward life must start with carefully chosen molecules in concentrations and purities far higher that what could have occurred naturally. They must also employ highly orchestrated experimental procedures (here, here, here, here). Any chain of chemical events on the early Earth that achieved the same results would also have been exceptionally rare.
Alternative Perspective
These challenges convince scientists who do not assume that life originated through undirected natural processes that the journey toward the first cell should be depicted very differently from that of Sutherland. Consider this figure which realistically presents what is necessary for life to arise:
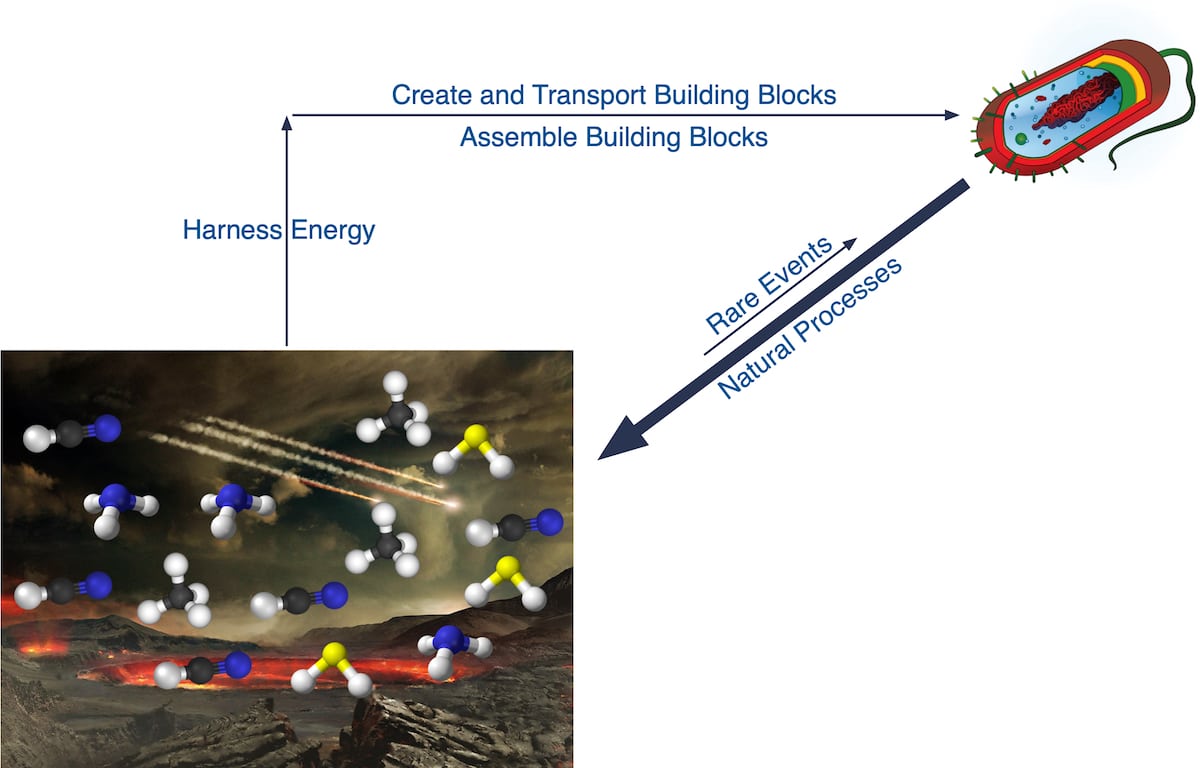
Figure 2 depicts four required processes for constructing a minimally complex cell:
- The building blocks of life must be synthesized, including proteins, RNA, DNA, and cellular structures.
- The building blocks must be transported to the same location. This step is far more difficult to explain through natural processes than most realize. The origin of the constituent molecules and linking them together require at least eight different environments.
- The building blocks must be properly assembled into a functional cell. The number of configurations corresponding to life divided by the number of all possible configurations is an unimaginably small ratio.
- Energy must be harnessed from the environment and converted into biologically useful forms, and that energy must be directed toward constructing a cell and then sustaining it. Continuous and reliable energy conversion and delivery can only be achieved with complex molecular machinery (here, here, here).
Figure 2 also depicts how every step toward life accomplished through a natural process corresponds to very “rare events.” In contrast, the natural processes continuously acting on an emerging protocell would have relentlessly driven it back toward biologically useless complex mixtures.
Even a 95 percent complete cell would irreversibly decompose into simpler molecules. Every chance step in the local environment toward cellular autonomy (e.g., two amino acids linking) would be matched by physical processes breaking apart the cell membrane, disassembling cellular machinery, and degrading proteins, RNA, and DNA. Every step forward would have been overshadowed by countless steps backward.
The Chicken or the Egg
Jeremy England elegantly describes the thermodynamic challenge for all life in his book Every Life Is on Fire: How Thermodynamics Explains the Origins of Living Things when he states:
Like all living things, plants are structures that absorb energy from specific sources in ways that lead to internal motions that correct or undo each incremental bit of falling to pieces that happens at every moment. This process goes on not only when a wound heals, but also much more instantaneously every time sunlight is used to regenerate a molecule of chemical fuel that was just burned up, or every time a molecular chaperone burns up some chemical fuel in order to help a protein that has become misfolded to get back into the correct, functional shape. The fuel-consuming, heat-dissipating activities of proofreading, quality control, and self-maintenance lie at the core of what living things are doing all the time to remain alive, and every one of these activities involves some kind of cyclical motion, whereby work absorbed from the environment perpetually drags things back up the mountain as each little downward slip and slide occurs.
The dilemma for life’s origin is that the machineries for “proofreading, quality control, and self-maintenance” are required early along the journey to life, but they can only be built by cells that are already fully autonomous. In other words, life cannot form unless life already exists.
Final Analysis
As I said in my previous article, I applaud Paul Rimmer’s civility, thoughtfulness, and stature as a committed theist who works in the field of origin of life research. Many might be attracted to his approach, and rightly so. I hope for the opportunity to have a dialogue with him someday. But just as hoping and believing in something doesn’t therefore make it true, being civil and thoughtful does not change the state of the scientific evidence. We must face reality, and when we realistically consider the complexity of biological systems, the quest to explain life’s origin through natural chemical processes shows every sign of being an impossible scientific goal.
In fact, those not committed to the philosophy of scientific materialism often see attempts to explain life’s origin through natural processes as falling into the same category as alchemy or the quest for a perpetual motion machine. In addition, the minimal requirements for cells, such as information processing, energy production, and error correction, represent indisputable evidence for direct design. The denial of design is not driven by the empirical data or theoretical analyses but by philosophical commitments.