 Evolution
Evolution
 Intelligent Design
Intelligent Design
Want to Harmonize Evolution and Design? Consider the Bacterial Flagellum
Our recent paper, “On the Relationship between Design and Evolution,” in the journal Religions, covers a lot of ground in response to Rope Kojonen’s book The Compatibility of Evolution and Design. Kojonen, a theologian, attempts to harmonize mainstream evolutionary theory with his version of design. One of the topics we consider is the bacterial flagellum. Kojonen believes the bacterial flagellum counts as evidence of design in a way that also dovetails with evolution. That is, he thinks that the flagellum is a powerful indicator of design and that it is also perfectly compatible with evolutionary theory. Thus, it poses no problem for his attempt to join “design” and “evolution” into one harmonious whole.
My co-authors — Stephen Dilley, Brian Miller, Emily Reeves — and I disagree with Kojonen about that and other things, as we have been relating in this series. We state in our article:
We …argue … that the type of design embodied in the bacterial flagellum is incompatible with mainstream evolutionary theory. This harms Kojonen’s philosophical attempt to harmonize ‘design’ and ‘evolution’. In effect, Kojonen’s conjunction of ‘evolution and design’ is at odds with itself: one of the conjuncts undermines the other. That is, the bacterial flagellum provides evidence of ‘design’ in a way that damages ‘evolution.’ [Emphasis in the original.]
To illuminate this problem, we start with an explanation of irreducible complexity:
An initial definition will start matters. In his 1996 book, Darwin’s Black Box, biochemist Michael Behe defines “irreducible complexity” as follows:
“In The Origin of Species Darwin stated: ‘If it could be demonstrated that any complex organ existed which could not possibly have been formed by numerous, successive, slight modifications, my theory would absolutely break down’. A system which meets Darwin’s criterion is one which exhibits irreducible complexity. By irreducible complexity I mean a single system composed of several well-matched, interacting parts that contribute to the basic function, wherein the removal of any one of the parts causes the system to effectively cease functioning.” (Behe 1996, p. 39)
Behe provides a mousetrap illustration that is helpful in this regard. He notes that a mousetrap is an irreducibly complex system: it is composed of five parts (a platform, spring, hammer, catch, and trigger), each of which is essential for the trap to work. A mousetrap with 80% of the parts does not catch 80% as many mice. Without a hammer, for example, the trap catches no mice. It simply does not work. All parts have to be in place for the trap to be functional.
A Positive Argument for Design
So Behe establishes why irreducible complexity is a problem for Darwinian explanations. But why is it best explained by intelligent design? We elaborate:
Behe’s broader argument is that IC systems are best explained by a mind rather than by mindless physical processes. We know from our repeated experience that intelligent agents create irreducibly complex systems — they can bring together an array of parts in order to achieve a particular outcome, whether that be a radio to catch soundwaves or a rocket ship to travel to space. By contrast, we know that stepwise physical processes, such as natural selection, must move towards immediate function (or else they wander stochastically, or perhaps even fail). For a process like natural selection to build some new feature, each step must confer a functional advantage that enhances the organism’s survival and reproduction. If not, the step is “blind” to natural selection, and there is no selective way to pass the trait on to the next generation. So, in human experience, gradual natural processes do not produce IC systems. But intelligent agents do. Thus, intelligent agency is a superior explanation for the origin of irreducibly complex systems.
“Divine Beauty” of the Bacterial Flagellum
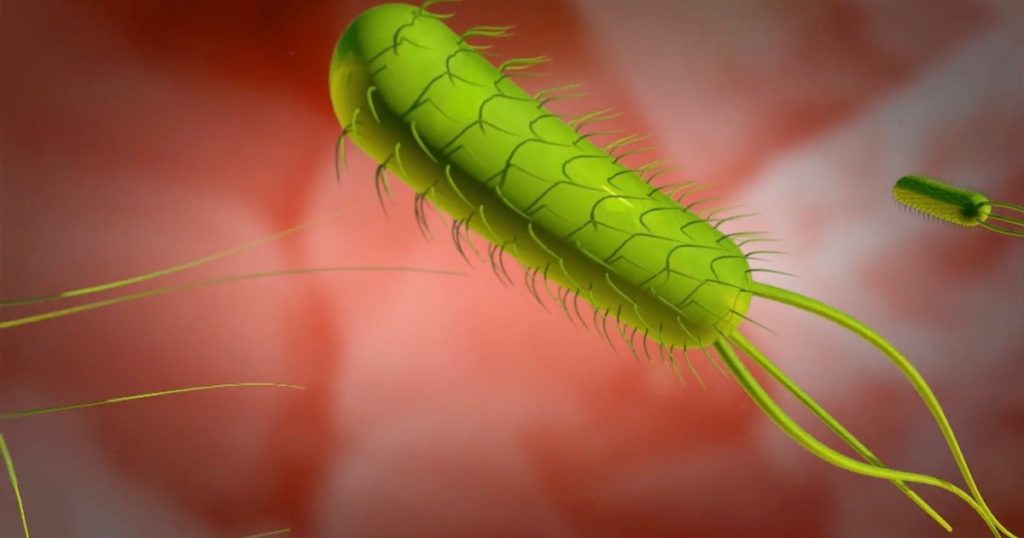
Of course, the most famous example of irreducible complexity is the bacterial flagellum. We introduce the flagellum and its impressive complexity:
Of the many examples of irreducible complexity that Behe cites, the most prominent is the bacterial flagellum. This organelle is a “true rotary engine” (Minnich and Meyer 2004) that propels a bacterium through liquid by rapidly spinning a whip-like tail. It operates much like an outboard motor.
[…]
Harvard biologist Howard Berg has deemed the bacterial flagellum “the most efficient machine in the universe” (quoted in Dembski 2004, p. 324). It is not difficult to see why. It is a true machine, having some 35–40 protein parts, each of which has an individual function, and which together perform an integrated function—complete with short-term memory, self-assembly, and an efficiency that surpasses human engineering. The flagellum has a highly efficient proton-powered rotary engine that operates at up to 100,000 rpm. It is one of the best understood molecular machines in science (Minnich and Meyer 2004).
5.2.2. The Engineering Logic of the Bacterial Flagellum
The engineering logic of the bacterial flagellum has been detailed by engineer and computer scientist Waldean Schulz (2021a, 2021b, 2021c). He has demonstrated that rotary propulsion requires several tightly integrated systems: a flagellar motor and a filament, part delivery mechanisms, an assembly process, and a navigation system. Each is essential for the flagellum to function, each is composed of multiple proteins, and each must meet very tight constraints dictated by an overarching design logic.
Subsystems of the flagellum include:
- A rotary engine with stators that generate greater torque in more viscous environments using an ingeniously engineered adaptive response strategy. The motor also has a clutch to suspend propulsion when doing so is advantageous (National Science Foundation 2008; Guttenplan et al. 2010).
- A propellor-like filament (Ikeda et al. 1996).
- Maintenance processes for the motor and filament (Schuhmacher et al. 2015; Ribardo et al. 2019; Zhuang and Lo 2020).
- An assembly process that incorporates an export gate which moves target proteins outside the bacterium in the right amount at the right time to construct the hook and filament (Waters et al. 2007; Minamino et al. 2014; Tan et al. 2021).
- Processes that suspend operations and eject flagellar proteins in threatening or low-nutrient environments as a form of risk management (Ferreira et al. 2019; Nedeljković et al. 2021).
- Chemotactic navigation that employs sensors which send a chemical signal to the motor to change direction when the bacterium is moving in a direction that is not favorable (Hamadeh et al. 2011; Othmer et al. 2013).
Numerous articles have been written on the ingenuity of each of these mechanisms, some of which have even served as models for human innovation (Mohammadi et al. 2017; Jiang et al. 2021; Tachiyama et al. 2022). One biologist observed that, “…the flagellum is so well designed and beautifully constructed by an ordered assembly pathway, even I, who am not a creationist, get an awe-inspiring feeling from its ‘divine’ beauty” (Aizawa 2009).
Problems for Darwinian Models
But does the bacterial flagellum pose a challenge to standard Darwinian explanations? We argue that it does — and this point is even recognized by Rope Kojonen, who is a thoughtful critic:
[B]oth comparative biology and knockout experiments on the flagellum show that it requires many coordinated protein parts in order to function (Macnab 1987; Pallen and Matzke 2006). A machine of this intricacy — in which all essential parts must be in place prior to any function — requires explanation. Kojonen sees the power of Behe’s argument:
“Since the core function of an irreducible complexity emerges only after all necessary parts are in place, it cannot plausibly be evolved in this direct way. After all, natural selection cannot select for a function that emerges only after all of the parts are in place, because selection cannot look to the future. Instead, the gradual evolution of the parts of a system like the flagellum would have to be favored by natural selection for some other reason, not because of increases in mobility.” (Kojonen 2021, pp. 116–17)
Moreover, Kojonen acknowledges that even the claim of “indirect” evolution of the bacterial flagellum faces severe challenges and would even require some measure of “serendipity”:
“Behe admits that an irreducibly complex system could in principle evolve in the tinkering, indirect fashion that Behe’s critics point to. However, he claims that, as the complexity of the system increases, the probability of such evolutionary accounts decreases. Because the proteins must fit together, the parts must be modified before serving in the new function. Thus, ‘analogous parts playing other roles in other systems cannot relieve the irreducible complexity of the new system; the focus simply shifts from “making” the components to “modifying” them’ (Behe 2006, pp. 112–13). Orr (1996), who is otherwise critical of Behe’s work, surprisingly agrees with this criticism: ‘we might think that some of the parts of an irreducibly complex system evolved step by step for some other purpose and were then recruited wholesale to a new function. But this is also unlikely. You may as well hope that half your car’s transmission will suddenly help out in the airbag department. Such things might happen very, very rarely, but they surely do not offer a general solution to irreducible complexity’. Here, the appeal to our common human experience of designing things supports the inference that creating complex teleological order is difficult. There is, indeed, quite a bit of serendipity in parts useful for one purpose being so easily adaptable to another role.” (Kojonen 2021, pp. 117–18)
Now Kojonen doesn’t consider the irreducible complexity of the flagellum to be a defeater for evolutionary models. Like many critics of Behe, he invokes co-option as a possible mechanism for how the flagellum evolved via an indirect evolutionary pathway. We’ll address that model of evolution in detail in a subsequent post, but for now it’s vital to appreciate the multi-subsystem complexity of the flagellum and how this poses a major challenge to its evolution:
Recall Kojonen’s earlier statement that the “crucial” issue concerning the evolution of proteins turns on “mutations”, including the number, timing, and rate needed to traverse from one functional protein to another. The challenge increases dramatically when assessing the origin of even one of the essential flagellar subsystems. For instance, undoubtedly, many mutations would be required for the ‘evolution’ of the flagellum’s navigation system (aka chemotaxis). To yield a functional system that provides an advantage to the organism, either all of these mutations would have to occur simultaneously — akin to a miracle — or each mutation (or set of mutations) would have to confer a functional advantage (or at least inflict no harm) at every step toward a fully operational system. Of particular note, the proteins composing the navigation system serve no other purpose in the bacterium, nor do they closely resemble any other proteins (see below). A partially materialized propulsion/navigation system, on the other hand, would not simply be “neutral” but rather disadvantageous to the cell given that the production of malformed proteins or some non-functional system would require energy to produce, yet would provide no offsetting benefit. In fact, because useless parts or proteins provide no advantage to the organism, they would likely be quickly degraded if not deleted (Gauger et al. 2010).
Again, in a follow-up post we’ll evaluate attempts to explain the evolution of the flagellum via co-option / exaptation / indirect evolution and show why they collide with many serious (even fatal) problems — not the least of which is the difficulty of evolving new proteins that Brian Miller has elaborated (see here, here, and here). The relevant citations can be found in our paper, here.
For now, I hope it’s clear that Kojonen’s effort to harmonize evolution and design (as he envisions it) is not so easy in the case of the bacterial flagellum. In effect, Kojonen wants to count the flagellum as designed, but also wants to ignore the fact that the type of design it displays — irreducible complexity — poses a major problem for evolution. Thus, he wants to join design and evolution, but only by setting aside some of the main features of the flagellum. This notably derails his attempted reconciliation.